




Содержание
Перейти к:
https://doi.org/10.29326/2304-196X-2026-15-2-164-169
Перейти к:
Введение. Вирусные респираторные заболевания являются серьезной проблемой для здоровья кошек во всем мире. Герпесвирус кошек 1-го типа (Feline herpesvirus type 1, FHV-1) – возбудитель высококонтагиозного заболевания, вызывающий инфекции верхних дыхательных путей, а также язвы конъюнктивы и роговицы. Герпесвирус 1-го типа считается основным этиологическим агентом респираторных инфекций у кошек, и, по оценкам специалистов, около 50–70% случаев респираторных инфекций связаны с данным возбудителем.
Цель исследования. Выделение вируса инфекционного ринотрахеита кошек из патматериала от больных животных в культуре клеток, идентификация и изучение его антигенных свойств.
Материалы и методы. Для выделения вируса in vitro использовали первично трипсинизированную культуру клеток почки эмбриона котенка, затем адаптировали его к перевиваемой культуре клеток почки кошки CRFK. Детекцию возбудителя инфекционного ринотрахеита кошек осуществляли с помощью полимеразной цепной реакции с обратной транскрипцией и полимеразной цепной реакции в режиме реального времени с применением специфичных праймеров, комплементарных участкам амплифицируемых фрагментов комплементарной ДНК. Вируснейтрализующие антитела в сыворотке крови иммунизированных кроликов определяли в реакции нейтрализации с использованием монослойной перевиваемой клеточной линии почки кошки CRFK.
Результаты. Установлено, что вирус инфекционного ринотрахеита кошек обнаруживается в конъюнктивальных, назальных, ротоглоточных смывах, реплицируется с проявлением выраженного цитопатического действия в первичной и субкультуре клеток почек эмбриона котенка и перевиваемой культуре клеток почки кошки CRFK, титр инфекционной активности через 72 ч после его инокуляции составлял (6,50 ± 0,25) и (7,40 ± 0,22) lg ТЦД50 /см3 соответственно.
Заключение. Полученный в результате исследований изолят герпесвируса кошек 1-го типа, обозначенный как штамм «Лавр», может использоваться для изготовления диагностических препаратов и средств специфической профилактики инфекционного ринотрахеита у кошек.
Галкина Т.С., Комарова А.А., Киселев А.М., Элизбарашвили Э.И. Выделение вируса инфекционного ринотрахеита кошек и его антигенные свойства. Ветеринария сегодня. 2026;15(2):164-169. https://doi.org/10.29326/2304-196X-2026-15-2-164-169
Galkina T.S., Komarova A.A., Kiselev A.M., Elizbarashvili E.I. Isolation of feline viral rhinotracheitis agent and its antigenic properties. Veterinary Science Today. 2026;15(2):164-169. (In Russ.) https://doi.org/10.29326/2304-196X-2026-15-2-164-169
Возбудитель инфекционного ринотрахеита кошек – ДНК-содержащий вирус, принадлежащий к семейству Orthoherpesviridae, виду Varicellovirus felidalpha 1 (Feline herpesvirus type 1, FHV-1, Felid alphaherpesvirus-1), являющийся причиной примерно 50–70% всех вирусных инфекций верхних дыхательных путей у домашних кошек [1][2][3][4][5][6]. Клинические признаки инфекции, вызванные FHV-1, обычно проявляются поражением глаз и дыхательной системы кошек. Характер и тяжесть заболевания могут различаться в зависимости от клинического случая. К основным распространенным симптомам инфекционного ринотрахеита кошек относятся конъюнктивит, стромальный кератит, секвестрация роговицы и сухой кератоконъюнктивит, серозные выделения из глаз и носа, чихание, кашель и анорексия [7][8][9]. Размножается вирус в эпителиальных клетках конъюнктивы и верхних дыхательных путей и поражает местные нейроны, устанавливая латентность в тройничном, крыловидно-небном и черепно-шейном ганглиях [10][11]. Хотя репликация вируса обычно ограничивается верхними дыхательными путями и конъюнктивой, во время острой фазы инфекции была выявлена виремия [12][13], также был описан случай негнойного менингоэнцефалита у кошек, это дает возможность полагать, что FHV-1 может вызывать более инвазивные заболевания [14]. Выздоровевшие кошки являются носителями вируса, у которых периодически случаются рецидивы, особенно после стресса [10]. Ранее было описано, что у кошек стресс, вызванный лактацией, может привести к реактивации латентной инфекции, опосредованной FHV-1, что в дальнейшем приводит к выделению вируса и возможной передаче инфекции. Заражению подвержены кошки любого возраста, пола и породы, но тяжелые клинические симптомы заболевания чаще встречаются у котят в возрасте от 2 до 6 мес., их смертность может достигать 50% [9][15].
Во многих исследованиях сообщалось о распространенности FHV-1 среди клинически здоровых и больных кошек. Частота выявления FHV-1 варьировалась в зависимости от места разведения, страны и клинического состояния кошек [15][16][17]. Исследования некоторых ученых показали, что более 90% кошек серопозитивны к FHV-1 и что минимум 80% остаются латентно инфицированными, а 45% выделяют вирус на протяжении всей жизни [4].
Все штаммы FHV-1 принадлежат к одной серогруппе, хотя для некоторых штаммов были зафиксированы незначительные генетические изменения [3][6][15][18]. В исследованиях R. Gaskell and K. Willoughby сообщалось о генетическом и антигенном сходстве FHV-1 с герпесвирусом собак 1-го типа (CHV-1), а также герпесвирусом тюленей 1-го типа (PhHV-1) и о перекрестной защите между FHV-1 и PhHV-1 [19].
Кошки с заболеваниями верхних дыхательных путей часто заражены одновременно FHV-1 и калицивирусом (FCV), что создает большие трудности при клинической диагностике, а также при выделении из биологического материала только FHV-1 в культуре клеток, поскольку большинство отобранных от кошек образцов являются коинфицированными [20][21][22][23]. Авторами многих исследований сообщалось об обнаружении коинфекции с использованием молекулярных методов, и выделение обоих вирусов в культуре клеток обычно использовалось для полной характеристики патогена [24].
С середины 1970-х гг. для борьбы с заболеванием, вызываемым FHV-1, используется вакцинация. В настоящее время в ветеринарной практике применяются как аттенуированные, так и инактивированные вакцины, которые хотя и могут снизить тяжесть течения заболевания инфекционного ринотрахеита у кошек, но не предотвращают заражения. Несмотря на широкое применение вакцин против вирусного ринотрахеита и калицивироза кошек в питомниках, заражение вирусами по-прежнему происходит часто, особенно при групповом содержании животных. Одна из возможных причин этого состоит в том, что даже вакцинированные кошки могут стать носителями FHV-1 после инфицирования и заразить восприимчивых животных при контакте. Другая причина может заключаться в том, что котята от инфицированных матерей могут заразиться и перенести герпесвирусную инфекцию в субклинической или легкой форме до вакцинации в возрасте 3–9 нед., когда уровень полученных от матери антител снижается [25][26][27].
В связи с высокой распространенностью инфекционного ринотрахеита в популяции кошек необходим постоянный эпизоотологический мониторинг. Исследования, направленные на выделение вируса, его идентификацию с целью дальнейшего изучения, обеспечат теоретическую основу для разработки новых средств специфической профилактики и борьбы с герпесвирусной инфекцией у кошек.
Образцы биоматериала были отобраны от кошек с подозрением на герпесвирусную инфекцию (24 пробы назальных, конъюнктивальных и ротоглоточных смывов) и поступили из ветеринарных клиник г. Владимира, Москвы, Вологды, Рыбинска, Нижнего Новгорода, а также от частных владельцев кошек в период с 2020 по 2021 г. Для подтверждения диагноза FHV-инфекции исследовали пробы биоматериала от больных кошек методами полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР) и полимеразной цепной реакции в режиме реального времени (ПЦР-РВ) при помощи коммерческих наборов «КАЛИЦИВИР» и «РИНОВИР» производства (ФБУН ЦНИИ Эпидемиологии Роспотребнадзора, Россия).
Проведение ПЦР. Для детекции ДНК возбудителя инфекционного ринотрахеита кошек проводили выделение нуклеиновой кислоты из биологического материала с помощью набора «РИБО-сорб» (ФБУН ЦНИИ Эпидемиологии Роспотребнадзора, Россия) в соответствии с инструкцией производителя, постановку ОТ-ПЦР и ПЦР-РВ выполняли с использованием специфичных праймеров, комплементарных участкам амплифицируемых фрагментов комплементарной ДНК. Учет результатов реакции амплификации осуществляли с применением технологии TaqMan (с использованием гибридизационно-флуоресцентной пробы) по значению цикла количественной оценки (Cq). В работе применяли олигонуклеотидные праймеры, представленные в таблице 1. Режимы термоциклирования отражены в таблице 2.
Таблица 1
Дизайн олигонуклеотидных праймеров для выявления ДНК возбудителя инфекционного ринотрахеита кошек с помощью ПЦР-РВ
Table 1
Design of oligonucleotide primers for detection of DNA of feline viral rhinotracheitis agent using qPCR
Название олигонуклеотида | 5’-3’-последовательность олигонуклеотида | Диапазон амплифицируемого фрагмента | Размер амплифицируемого фрагмента |
FHV-F1-66684 | AGATTTGCCGCACCATACCTTC | 66684…67221 | 538 п. н. |
FHV-R1-67199 | GATCTCCATTTTGGTCGGAGAGC |
Таблица 2
Режимы термоциклирования при детекции ДНК возбудителя инфекционного ринотрахеита кошек в ПЦР-РВ
Table 2
Thermocycling conditions for detection of DNA of feline viral rhinotracheitis agent using qPCR
Название этапа | Название подэтапа | Температура, °С | Время | Количество циклов |
Предварительная денатурация | – | 98 | 3 мин | 1 |
ПЦР-РВ | денатурация | 95 | 15 с | 40 |
отжиг праймеров | 60 | 30 с | ||
элонгация* | 72 | 30 с |
* детектирование сигнала проводится на этапе элонгации на канале Green. Термоциклирование проходило в течение ≈ 1,5 ч (signal detection is carried out during the elongation step on the Green channel. Thermocycling was run for about 1.5 hours).
Приготовление вирусной суспензии для заражения культур клеток. Образцы со слизистой оболочки рта и носа, а также конъюнктивы были получены с помощью стерильных ватных тампонов, которые помещали в фосфатно-солевой буфер (PBS), затем центрифугировали при 3000 об/мин в течение 10 мин, пропускали через фильтры с диаметром пор 0,22 мкм и выдерживали при минус 80 °C до использования. Полученные таким образом пробы применяли для заражения культур клеток.
Выделение FHV-1. Для выделения вируса in vitro использовалась первично трипсинизированная и субкультура клеток почки эмбриона котенка, которую выращивали в питательной среде Dulbecco’s Modified Eagle Medium DMEM (ООО «БиолоТ», Россия) с добавлением 10% фетальной сыворотки крупного рогатого скота (HyClone Laboratories LLC, США), 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина. Клетки выращивали в стандартных условиях, используемых для культивирования: при температуре (37,0 ± 0,5) °С в атмосфере 5% CO2 и при влажности воздуха 90%. Вирусные суспензии, приготовленные из проб биоматериала, положительные на FHV-1 при исследовании посредством ПЦР-РВ, вносили в объеме 500 мкл в монослой культуры клеток во флаконы с площадью роста 25 см³ (TPP, Швейцария) и инкубировали, как указано выше (стандартные условия). После завершения периода адсорбции продолжительностью в 60 мин добавляли среду DMEM с 1% фетальной сыворотки крови телят. Инфицированную культуру клеток культивировали в течение 5 сут. Всего проводили не менее 5 «слепых» пассажей до выявления цитопатогенного действия (ЦПД) вируса в культуре клеток. Если ЦПД не наблюдалось на протяжении 5 пассажей, образец считался отрицательным. В случае если ЦПД появлялось, клетки замораживали и размораживали три раза для высвобождения вируса. В последующей работе вирус адаптировали и культивировали в перевиваемой линии клеток почки кошки CRFK (Crandell-Rees Feline Kidney), которую выращивали в питательной среде DMEM с добавлением 10% фетальной сыворотки крупного рогатого скота (HyClone Laboratories LLC, США), 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина и инкубировали, как указано выше (стандартные условия).
Определение титра инфекционной активности FHV-1. Титр инфекционной активности FHV-1 определяли с помощью клеток линии CRFK в 96-луночных планшетах в соотношении 1,5 × 10⁵ кл/мл с использованием 100 мкл на лунку и культивировали, как указано выше, в течение 24 ч. Добавляли разведения FHV-1 и оставляли на контакте в течение 60 мин, затем надосадочную жидкость удаляли и заменяли 200 мкл DMEM, содержащей 1% фетальной сыворотки крови телят. Наблюдение за клетками проводили каждые 12 ч в течение 5 дней и рассчитывали 50%-ю инфекционную дозу для культуры клеток (TЦД50) с использованием метода Рида – Менча.
Определение влияния множественности заражения на инфекционную активность FHV-1. Для изучения влияния множественности заражения (MOI) на инфекционную активность FHV-1 применяли дозы заражения 0,0001; 0,001; 0,01 и 0,1 ТЦД50/кл in vitro. Вирус инокулировали в перевиваемую линию клеток почки кошки CRFK в дозах 0,1; 0,01; 0,001 и 0,0001 МОI соответственно. Культуру клеток собирали через 12, 24, 36, 48 и 72 ч после инокуляции. Значение TЦД50 определяли с помощью метода Рида – Менча.
Животные. Для оценки антигенных свойств выделенного изолята FHV-1 использовались кролики 45-суточного возраста с массой тела 1,5–3,0 кг (n = 4), содержащиеся в лабораторно-виварном корпусе ФГБУ «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ», г. Владимир).
Все процедуры, выполненные с участием животных, соответствовали этическим стандартам, принятым Европейской конвенцией ETS № 123, и одобрены Комиссией по биоэтике ФГБУ «ВНИИЗЖ».
Постановка реакции нейтрализации (VN, РН). Нейтрализующие FHV-1 антитела определяли с помощью РН в монослойной перевиваемой клеточной линии почки кошки CRFK. Титр антител устанавливали путем последовательного разведения образца сыворотки крови иммунизированных кроликов, который затем добавлялся к стандартному количеству вируса. Краткое описание постановки реакции: 50 мкл разведенной сыворотки крови кроликов и 50 мкл инфекционной культуральной среды, содержащей 100 TЦД50 выбранного штамма FHV-1, смешивали, инкубировали в течение 2 ч при (37,0 ± 0,5) °С и 5% CO2. По истечении времени инкубации смесь «вирус – антитело» инокулировали в клеточные культуры CRFK в 96-луночные микропланшеты с поверхностью дна CellBIND. Каждое разведение сыворотки крови кроликов тестировали с использованием четырех лунок на разведение. Культуры инкубировали в течение 5 дней при (37,0 ± 0,5) °С и 5% CO2. Реакцию учитывали визуально при помощи инвертированного микроскопа Olympus CKX53 (Olympus Corporation, Япония). Титр вируснейтрализующих антител определяли как величину, обратную наибольшему разведению, которое предотвращало инфицирование клеток.
Статистический анализ результатов. Обработка полученных данных производилась с использованием статистических методов в программе Microsoft Exсel. Определяли среднегрупповые значения титров и стандартное отклонение. Расчет титра вируснейтрализующих антител в сыворотках крови кроликов проводили по формуле Кербера и выражали в логарифмах с основанием 2 (log2).
Пробы биоматериала от больных кошек, отобранные для выделения вируса, были исследованы с помощью ОТ-ПЦР с горизонтальным гель-электрофорезом для выявления наличия генома FHV-1 и ПЦР-РВ для количественной оценки ДНК FHV-1.
Детекцию результатов на FHV-1 осуществляли, регистрируя значения циклов количественной оценки ДНК исследуемых образцов (рис. 1, табл. 3).
Рис. 1. Сигмоиды накопления сигналов флуоресценции при проведении ПЦР-РВ для выявления ДНК FHV-1
Fig. 1. Sigmoid curves of fluorescence signal accumulation during qPCR for the detection of FHV-1 DNA
Таблица 3
Результаты исследования проб на выявление генома возбудителя инфекционного ринотрахеита кошек
Table 3
Results of testing the samples for the detection of the genome of feline viral rhinotracheitis agent
№ п/п | Изоляты | Значение цикла количественной оценки (Cq) | № п/п | Изоляты | Значение цикла количественной оценки (Cq) |
1 | К-03/20 | 23,63 | 13 | В-07/20 | 25,09 |
2 | КВ-03/20 | 23,76 | 14 | М-04/21 | 26,67 |
3 | КВ-05/20 | 23,90 | 15 | М-10/20 | 26,52 |
4 | В-07/21 | 23,16 | 16 | В-11/21 | 25,69 |
5 | К-04/21 | 23,64 | 17 | ВЛ-09/20 | 24,97 |
6 | ВЛ-05/21 | 23,62 | 18 | М-07/20 | 23,81 |
7 | М-06/20 | 23,36 | 19 | М-05/21 | 25,12 |
8 | НН-09/21 | 24,14 | 20 | Рнн-12/21 | 24,94 |
9 | НО-10/21 | 23,45 | 21 | ВР-06/21 | 26,31 |
10 | Р-11/20 | 24,90 | 22 | В-02/20 | 25,00 |
11 | МО-08/21 | 24,17 | 23 | И-04/21 | 24,42 |
12 | Р-06/21 | 23,64 | 24 | МО-06/20 | 26,15 |
С помощью ПЦР-тестов геном FHV-1 был обнаружен во всех 24 (100%) пробах, количество копий ДНК возбудителя инфекционного ринотрахеита кошек во всех пробах было примерно одинаковое. Попарный сравнительный анализ показал, что нуклеотидные последовательности выявленных 24 изолятов FHV-1 гомологичны на 99–100%.
В ходе исследований методом ПЦР установлено, что все 24 пробы содержат нуклеиновую кислоту FHV-1, из них в 19 случаях (№ 1–3, 5, 6, 8, 10–14, 16–23) наблюдали коинфицирование FCV и FHV-1; в пробах № 4, 7, 9, 15 и 24 был обнаружен только геном FHV-1, данные образцы были использованы в дальнейшей работе.
Как видно из таблицы 3, наименьшее накопление ДНК FHV-1 – в пробах № 1–3, 5, 6, 8, 10–14, 16–23; наибольшее – в пробе № 4.
Вирусные суспензии, приготовленные из проб № 4, 7, 9, 15 и 24 (изоляты В-07/21, М-06/20, НО-10/21, М-10/20 и МО-06/20 соответственно), вносили в объеме 500 мкл в сформированный монослой первично трипсинизированной культуры клеток почки эмбриона котенка во флаконы с площадью роста 25 см³.
В результате нескольких пассажей в первично трипсинизированной и субкультуре клеток почки эмбриона котенка были выделены цитопатогенные изоляты FHV-1 (В-07/21, М-06/20, НО-10/21), которые накапливались после 3 пассажей в титрах инфекционной активности (6,50 ± 0,25), (3,91 ± 0,14) и (2,75 ± 0,43) lg ТДЦ50/см³ соответственно. У изолятов М-10/20 и МО-06/20 FHV-1 в культурах клеток почки эмбриона котенка инфекционный титр был на уровне (1,66 ± 0,38) lg ТДЦ50/см³ в течение 2 пассажей и к 3-му пассажу ЦПД не наблюдалось, поэтому данные материалы в дальнейшем не использовались в работе.
В связи с тем что нуклеотидные последовательности цитопатогенных изолятов FHV-1 гомологичны на 99–100% (по результатам попарного сравнительного анализа), для дальнейших исследований был выбран изолят В-07/21 с наиболее высоким инфекционным титром в первично трипсинизированной и субкультуре клеток почки эмбриона котенка. Изолят В-07/21 FHV-1 адаптировали и культивировали в перевиваемой линии клеток почки кошки CRFK. Первые признаки характерного ЦПД отмечали через 24 ч после заражения монослоя, которое выражалось в набухании и округлении клеток, при сохранении целостности монослоя. Через 48–72 ч после инфицирования монослоя наблюдали цитогамию, образование отдельных очагов больших округлых клеток, в конечном итоге они полностью отделялись и оставались во взвешенном состоянии через 96 ч.
На рисунках 2A и 2B видно, что выделенный изолят В-07/21 FHV-1 вызывал выраженное ЦПД в перевиваемой культуре клеток CRFK через 72 ч после инфицирования. Вирус при этом накапливался после 3 пассажей в титрах инфекционной активности (7,33 ± 0,14) lg ТДЦ50/см³.

Рис. 2. Цитопатический эффект изолята В-07/21 FHV-1 в клетках CRFK через 72 ч после инфицирования: A – увеличение 4×; B – увеличение 10×; С – контрольные (неинфицированные) клетки CRFK (увеличение 4×)
Fig. 2. Cytopathic effect of V-07/21 FHV-1 isolate in CRFK cells 72 hours after infection: A – magnification 4×; B – magnification 10×; C – control (non-infected) CRFK cells (magnification 4×)
Результаты изучения влияния множественности заражения на инфекционную активность В-07/21 FHV-1 в культуре клеток CRFK показали, что чем ниже доза введения инокулята, тем медленнее реплицируется вирус. Самый высокий титр инфекционной активности В-07/21 FHV-1 зарегистрирован через 72 ч после инокуляции в дозе 0,01 МОI, который составил (7,40 ± 0,22) lg ТЦД50/см³ (рис. 3).
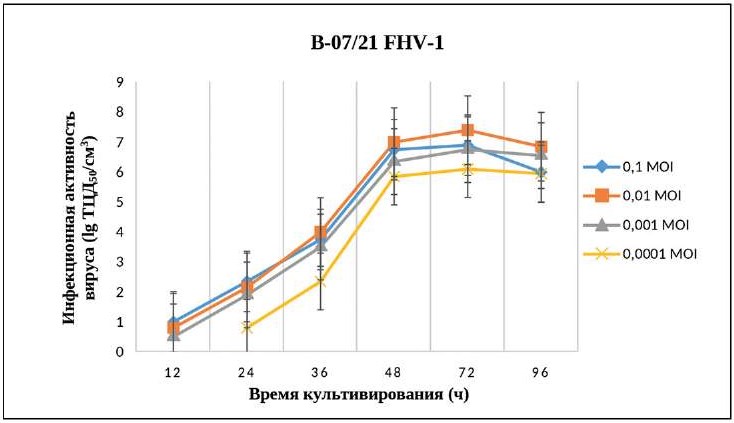
Рис. 3. Способность к репликации изолята В-07/21 FHV-1 при различных дозах инокуляции и времени культивирования в культуре клеток CRFK (n = 5, p < 0,05)
Fig. 3. Replication capacity of FHV-1 isolate V-07/21 at various inoculation doses and cultivation times in CRFK cell culture (n = 5, p < 0.05)
Для изучения антигенных свойств изолята В-07/21 FHV-1 проводили иммунизацию 4 кроликов инактивированным материалом в дозе 1,0 см³ двукратно с интервалом 21 сут внутримышечно. Титр антител к антигену вируса инфекционного ринотрахеита кошек в сыворотках крови кроликов определяли в РН до введения и через 21 и 35 сут после иммунизации. Кролики до введения инактивированного материала были серонегативны к вирусу FHV-1.
Результаты исследования антигенной активности изолята В-07/21 FHV-1 на кроликах представлены на рисунке 4. Титр вируснейтрализующих антител к FHV-1 у иммунизированных кроликов после двукратного введения инактивированного вируссодержащего материала на 21-е сут составил в среднем по группе (4,91 ± 0,15) log2 SN50, на 35-е сутки титр антител в сыворотке крови достигал статистически значимого показателя, составляющего (6,87 ± 0,59) log2 SN50 (n = 3, p < 0,05), что свидетельствует о высокой антигенной активности вируса.
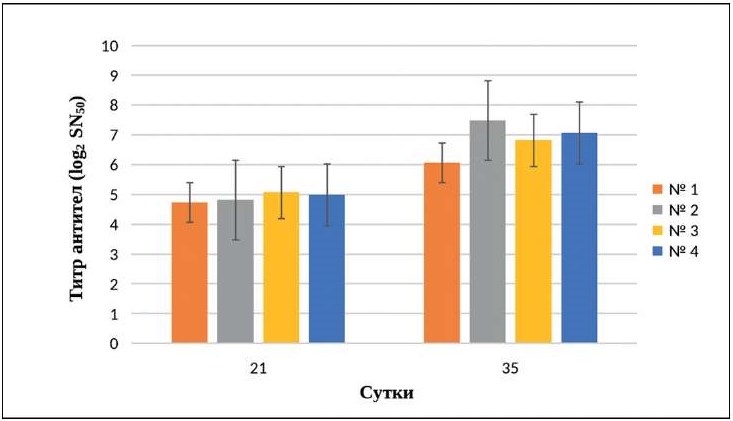
Рис. 4. Динамика сероконверсии к В-07/21 FHV-1 у кроликов, по данным РН (n = 3, p < 0,05)
Fig. 4. Dynamics of seroconversion to FHV-1 V-07/21 in rabbits, according to VN test results (n = 3, p < 0.05)
Выделенный изолят В-07/21 FHV-1 обозначили как штамм «Лавр», который был депонирован в качестве производственно-контрольного штамма во Всероссийскую государственную коллекцию экзотических типов вируса ящура и других патогенов животных ФГБУ «ВНИИЗЖ» под регистрационным номером № 457-деп/23-7-ГКШМ ФГБУ «ВНИИЗЖ». На штамм был получен патент № 2806603 C1 «Штамм “Лавр” вируса Alphaherpesvirus 1 инфекционного ринотрахеита кошек для изготовления биопрепаратов для диагностики и специфической профилактики инфекционного ринотрахеита кошек»1.
Анализ полученных результатов исследований позволяет сделать заключение, что вирус инфекционного ринотрахеита кошек обнаруживается в конъюнктивальных, назальных, ротоглоточных смывах. Установлено, что к герпесвирусу 1-го типа в равной степени чувствительны первичные и субкультуры клеток почек эмбриона котенка, перевиваемая культура клеток почки кошки CRFK, где он реплицируется с проявлением выраженного ЦПД. Однако, на наш взгляд, наиболее перспективной культурой клеток для репродукции возбудителя инфекционного ринотрахеита кошек является перевиваемая культура клеток почки кошки CRFK. Использование первично трипсинизированной культуры и субкультуры почек эмбриона котенка для последующего культивирования вируса затруднена в связи с их низкой технологичностью и из-за постоянной потребности в донорах органов. Полученные результаты исследований могут использоваться в диагностических целях и при разработке специфических средств профилактики инфекционного ринотрахеита у кошек.
Вклад авторов: Галкина Т. С. – идея и дизайн исследования, проведение экспериментов, систематизация результатов, анализ литературы, написание текста, утверждение окончательного варианта статьи; Комарова А. А. – вирусологические исследования, обработка биологического материала, анализ и интерпретация данных; Киселев А. М. – вирусологические исследования, обработка биологического материала; Элизбарашвили Э. И. – научная консультация.
Contribution of the authors: Galkina T. S. – study concept and design, experimental work, data organization, literature review, manuscript writing, and final approval of the article; Komarova A. A. – virological studies, biological sample processing, data analysis and interpretation; Kiselev A. M. – virological studies, biological sample processing; Elizbarashvili E. I. – scientific consultation.
1. https://patentimages.storage.googleapis.com/7e/6d/f6/7c51d22d54701f/RU2806603C1.pdf
1. Рахманина М. М., Рахманина Н. А., Элизбарашвили Э. И., Уласов В. И., Сулимов А. А. Разработка биопрепаратов для диагностики, профилактики и лечения при вирусных болезнях кошек. Ветеринария. 2011; (1): 48–51. https://elibrary.ru/ncxedt Rahmanina M. M., Rahmanina N. A., Elizbarashvili E. I., Ulasov V. I., Sulimov A. A. Creation of biological products for diagnostics, preventive maintenance and processings at feline viral infections. Veterinariya. 2011; (1): 48–51. https://elibrary.ru/ncxedt (in Russ.)
2. Кокорина Е. Г., Элизбарашвили Э. И. Сравнительная оценка репродуктивных свойств вируса инфекционного ринотрахеита кошек на различных культурах клеток. Ветеринария сегодня. 2018; (3): 69–72. https://doi.org/10.29326/2304-196X-2018-3-26-69-72 Kokorina Ye. G., Elizbarashvili E. I. Comparative studies of feline viral rhinotracheitis virus for its replication properties in different cell cultures. Veterinary Science Today. 2018; (3): 69–72. https://doi.org/10.29326/2304-196X-2018-3-26-69-72
3. Maes R. Felid herpesvirus type 1 infection in cats: a natural host model for alphaherpesvirus pathogenesis. International Scholarly Research Notices. 2012; 2012 (1):495830. https://doi.org/10.5402/2012/495830
4. Gaskell R., Dawson S., Radford A., Thiry E. Feline herpesvirus. Veterinary Research. 2007; 38 (2): 337–354. https://doi.org/10.1051/vetres:2006063
5. Davison A. J., Eberle R., Ehlers B., Hayward G. S., McGeoch D. J., Minson A. C., et al. The order Herpesvirales. Archives of Virology. 2009; 154 (1): 171–177. https://doi.org/10.1007/s00705-008-0278-4
6. Crandell R. A., Maurer F. D. Isolation of a feline virus associated with intranuclear inclusion bodies. Proceedings of the Society of Experimental Biology and Medicine. 1958; 97 (3): 487–490. https://doi.org/10.3181/00379727-97-23783
7. Stiles J. Treatment of cats with ocular disease attributable to herpesvirus infection: 17 cases (1983–1993). Journal of the American Veterinary Medical Association. 1995; 207 (5): 599–603. https://pubmed.ncbi.nlm.nih.gov/7649774
8. Sun H., Li Y., Jiao W., Liu C., Liu X., Wang H., et al. Isolation and identification of feline herpesvirus type 1 from a South China tiger in China. Viruses. 2014; 6 (3): 1004–1014. https://doi.org/10.3390/v6031004
9. Marino M. E., Mironovich M. A., Inek N. E., Citino S. B., Emerson J. A., Maggs D. J. et al. Full viral genome sequencing and phylogenomic analysis of feline herpesvirus type 1 (FHV-1) in cheetahs (Acinonyx jubatus). Viruses. 2021; 13 (11):2307. https://doi.org/10.3390/v13112307
10. Yang J., Mei R., Sun K., Xu F., Jia F. Detection of neutralizing antibody to Feline herpesvirus type 1 in cat serum. Comparative Immunology, Microbiology and Infectious Diseases. 2025; 124:102419. https://doi.org/10.1016/j.cimid.2025.102419
11. Townsend W. M., Jacobi S., Tai S.-H., Kiupel M., Wise A. G., Maes R. K. Ocular and neural distribution of feline herpesvirus-1 during active and latent experimental infection in cats. BMC Veterinary Research. 2013; 9:185. https://doi.org/10.1186/1746-6148-9-185
12. Swenson C. L., Gardner K., Arnoczky S. P. Infectious feline herpesvirus detected in distant bone and tendon following mucosal inoculation of specific pathogen-free cats. Veterinary Microbiology. 2012; 160 (3–4): 484–487. https://doi.org/10.1016/j.vetmic.2012.06.028
13. Westermeyer H. D., Thomasy S. M., Kado-Fong H., Maggs D. J. Assessment of viremia associated with experimental primary feline herpesvirus infection or presumed herpetic recrudescence in cats. American Journal of Veterinary Research. 2009; 70 (1): 99–104. https://doi.org/10.2460/ajvr.70.1.99
14. Hora A. S., Tonietti P. O., Guerra J. M., Leme M. C., Pena H. F. J., Maiorka P. C., Brandão P. E. Felid herpesvirus 1 as a causative agent of severe nonsuppurative meningoencephalitis in a domestic cat. Journal of Clinical Microbiology. 2013; 51 (2): 676–679. https://doi.org/10.1128/jcm.02462-12
15. Kang B.-T., Park H.-M. Prevalence of feline herpesvirus 1, feline calicivirus and Chlamydophila felis in clinically normal cats at a Korean animal shelter. Journal of Veterinary Science. 2008; 9 (2): 207–209. https://doi.org/10.4142/jvs.2008.9.2.207
16. Magouz A., Lokman M. S., Albrakati A., Elmahallawy E. K. First report of isolation and molecular characterization of felid herpesvirus-1 from symptomatic domestic cats in Egypt. Veterinary Sciences. 2022; 9 (2):81. https://doi.org/10.3390/vetsci9020081
17. Holst B. S., Berndtsson L. T., Englund L. Isolation of feline herpesvirus-1 and feline calicivirus from healthy cats in Swedish breeding catteries. Journal of Feline Medicine and Surgery. 2005; 7 (6): 325–331. https://doi.org/10.1016/j.jfms.2005.03.002
18. Vaz P. K., Job N., Horsington J., Ficorilli N., Studdert M. J., Hartley C. A., et al. Low genetic diversity among historical and contemporary clinical isolates of felid herpesvirus 1. BMC Genomics. 2016; 17:704. https://doi.org/10.1186/s12864-016-3050-2
19. Gaskell R., Willoughby K. Herpesviruses of carnivores. Veterinary Microbiology. 1999; 69 (1–2): 73–88. https://doi.org/10.1016/S0378-1135(99)00092-9
20. Schulz C., Hartmann K., Mueller R. S., Helps C., Schulz B. S. Sampling sites for detection of feline herpesvirus-1, feline calicivirus and Chlamydia felis in cats with feline upper respiratory tract disease. Journal of Feline Medicine and Surgery. 2015; 17 (12): 1012–1019. https://doi.org/10.1177/1098612X15569615
21. Walter J., Foley P., Yason C., Vanderstichel R., Muckle A. Prevalence of feline herpesvirus-1, feline calicivirus, Chlamydia felis, and Bordetella bronchiseptica in a population of shelter cats on Prince Edward Island. Canadian Journal of Veterinary Research. 2020; 84 (3): 181–188. https://pmc.ncbi.nlm.nih.gov/articles/PMC7301681
22. Guo H., Miao Q., Zhu J., Yang Z., Liu G. Isolation and molecular characterization of a virulent systemic feline calicivirus isolated in China. Infection, Genetics and Evolution. 2018; 65: 425–429. https://doi.org/10.1016/j.meegid.2018.08.029
23. Zheng H., Yue H., Wang B., Yu X., Liu Y., Yu J., et al. An efficient method for the selective isolation of feline herpesvirus 1(FHV-1) in feline calicivirus (FCV) coinfected specimens. BMC Veterinary Research. 2025; 21:321. https://doi.org/10.1186/s12917-025-04786-w
24. Litster A., Wu C. C., Leutenegger C. M. Detection of feline upper respiratory tract disease pathogens using a commercially available real-time PCR test. The Veterinary Journal. 2015; 206 (2): 149–153. https://doi.org/10.1016/j.tvjl.2015.08.001
25. Jas D., Aeberlé C., Lacombe V., Guiot A. L., Poulet H. Onset of immunity in kittens after vaccination with a non-adjuvanted vaccine against feline panleucopenia, feline calicivirus and feline herpesvirus. The Veterinary Journal. 2008; 182 (1): 86–93. https://doi.org/10.1016/j.tvjl.2008.05.025
26. Lappin M. R., Sebring R. W., Porter M., Radecki S. J., Veir J. Effects of a single dose of an intranasal feline herpesvirus 1, calicivirus, and panleukopenia vaccine on clinical signs and virus shedding after challenge with virulent feline herpesvirus 1. Journal of Feline Medicine and Surgery. 2006; 8 (3): 158–163. https://doi.org/10.1016/j.jfms.2005.12.001
27. Slater E., York C. Comparative studies on parenteral and intranasal inoculation of an attenuated feline herpes virus. Developments in Biological Standardization. 1976; 33: 410–416. https://pubmed.ncbi.nlm.nih.gov/182601
Галкина Татьяна Сергеевна, канд. вет. наук, заведующий лабораторией профилактики болезней мелких домашних животных
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Комарова Анна Александровна, ведущий ветеринарный врач лаборатории профилактики болезней мелких домашних животных
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Киселев Алексей Максимович, канд. вет. наук, ветеринарный врач лаборатории профилактики болезней мелких домашних животных
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Элизбарашвили Элизбар Иосифович, канд. вет. наук,
главный специалист отдела вирусологии
Звенигородское шоссе, 5, г. Москва, 123022
Галкина Т.С., Комарова А.А., Киселев А.М., Элизбарашвили Э.И. Выделение вируса инфекционного ринотрахеита кошек и его антигенные свойства. Ветеринария сегодня. 2026;15(2):164-169. https://doi.org/10.29326/2304-196X-2026-15-2-164-169
Galkina T.S., Komarova A.A., Kiselev A.M., Elizbarashvili E.I. Isolation of feline viral rhinotracheitis agent and its antigenic properties. Veterinary Science Today. 2026;15(2):164-169. (In Russ.) https://doi.org/10.29326/2304-196X-2026-15-2-164-169
600901, Владимирская область, город Владимир, микрорайон Юрьевец, улица Гвардейская, дом 6
ФГБУ «ВНИИЗЖ»
тел.: 8 (4922) 26-15-12, доб. 22-27
E-mail: nikeshina@arriah.ru






























