




Содержание
Перейти к:
https://doi.org/10.29326/2304-196X-2026-15-2-110-122
Перейти к:
Введение. Вирусный гепатит утят –высококонтагиозное острое заболевание молодняка уток, наносящее значительный экономический ущерб утководческим хозяйствам и входящее в перечень нотифицируемых болезней Всемирной организации здравоохранения животных. Наиболее распространены три типа вируса, из которых тип 1 встречается повсеместно и является основным этиологическим агентом в большинстве регионов мира.
Цель исследования. Систематизация и анализ современных подходов к профилактике вирусного гепатита утят, обзор перспективных стратегий разработки рекомбинантных и поливалентных вакцин.
Материалы и методы. Поиск и анализ литературных источников проводился с использованием баз данных Scopus, Web of Science, Google Scholar, PubMed, ScienceDirect и РИНЦ.
Результаты. В статье приведен анализ современной эпизоотической ситуации по вирусному гепатиту утят, рассмотрены особенности патогенеза, иммунного ответа и диагностики заболевания. Представлены сравнительные данные по характеристикам различных вакцин против вирусного гепатита утят: их способу производства, эффективности, схемам иммунизации, формированию иммунитета, преимуществам и недостаткам, наличию на коммерческом рынке. Обобщены существующие подходы к защите поголовья и предложены направления дальнейших исследований.
Заключение. Несмотря на наличие большого количества разнообразных зарубежных вакцин против вирусного гепатита утят, в Российской Федерации единственной производимой вакциной является вакцина против вирусного гепатита утят из штамма «ВГНКИ-К» эмбриональная (ФГБУ «ВНИИЗЖ», Россия), которая имеет достаточно ограниченный срок годности (9 мес.). В связи с этим в ФГБУ «ВНИИЗЖ» проводится научно-исследовательская работа по разработке живой лиофилизированной вакцины с увеличенным сроком годности.
Куникова Е.Д., Волков М.С., Мороз Н.В. Специфическая профилактика вирусного гепатита утят (обзор). Ветеринария сегодня. 2026;15(2):110-122. https://doi.org/10.29326/2304-196X-2026-15-2-110-122
Kunikova E.D., Volkov M.S., Moroz N.V. Specific prevention of duck virus hepatitis (review). Veterinary Science Today. 2026;15(2):110-122. (In Russ.) https://doi.org/10.29326/2304-196X-2026-15-2-110-122
Вирусный гепатит утят (ВГУ) – высококонтагиозное остропротекающее заболевание, к которому восприимчивы утята до 3–4-недельного возраста, характеризующееся поражением печени и геморрагическим диатезом, сопровождающееся угнетением, желтухой и высокой смертностью среди молодняка [1][2][3]. Различают три типа возбудителя ВГУ. Вирус типа 1 (DHAV тип I, или DHV-1) относится к роду Avihepatovirus, семейству Picornaviridae, включает 3 серотипа (DHAV-1, или DHAV-A; DHAV-2, или DHAV-B; DHAV-3, или DHAV-C). Возбудители типов 2 и 3 (DHAV тип II, или DHV-2; DHAV тип III, или DHV-3) являются представителями семейства Astroviridae: астровирус уток типа 1 (DAstV-1) и астровирус уток типа 2 (DAstV-2) соответственно [4][5]. Вирус 2-го типа был выделен в Англии и 3-го – в США [6].
Актуальность изучения ВГУ обусловлена тяжелым течением заболевания у молодняка, высокой скоростью его распространения и значительным экономическим ущербом, который проявляется в виде потерь не только вследствие высокой летальности среди утят, но и отставания в росте и развитии переболевших особей, снижения продуктивности взрослого поголовья, а также дополнительных затрат на проведение ограничительных мероприятий [2].
Целью настоящего обзора является систематизация современных данных о ВГУ с акцентом на вопросы специфической профилактики, в частности вакцинопрофилактики, а также анализ ситуации в Российской Федерации, где используется лишь одна зарегистрированная вакцина отечественного производства.
Вирусный гепатит утят впервые был официально зарегистрирован в 1949 г. в США, после чего заболевание стало быстро распространяться в странах с развитым утководством: в Канаде, Бразилии, Германии, Франции, Украине, Англии, Бельгии, Индии, Чехии, Японии, Польше. Летальность при ВГУ колебалась от 30 до 95% [7][8]. В 1958 г. в СССР на территории Белгородской и Харьковской областей была зафиксирована первая вспышка ВГУ [5].
Было описано три различных типа возбудителя ВГУ, и все они первоначально были классифицированы как пикорнавирусы, причем тип 1 считался наиболее похожим на вирусы рода Enterovirus. Вирусы гепатита утят 2-го и 3-го типов были впоследствии отнесены к семейству Astroviridae и переименованы в астровирусы уток. Из всех трех типов возбудитель ВГУ типа 1 является наиболее широко распространенным и наиболее вирулентным [9].
Серотип 1 (DHAV-1) циркулирует во многих регионах мира, таких как Англия, Польша, Китай и Египет [10]. Серотип 2 (DHAV-2), напротив, встречается только на Тайване и в Китае [9]. Серотип 3 (DHAV-3) – это новый серотип, который появился в Корее в 2003 г., а затем распространился в Китае, Вьетнаме и недавно – в Египте (рис. 1) [10][11][12][13].
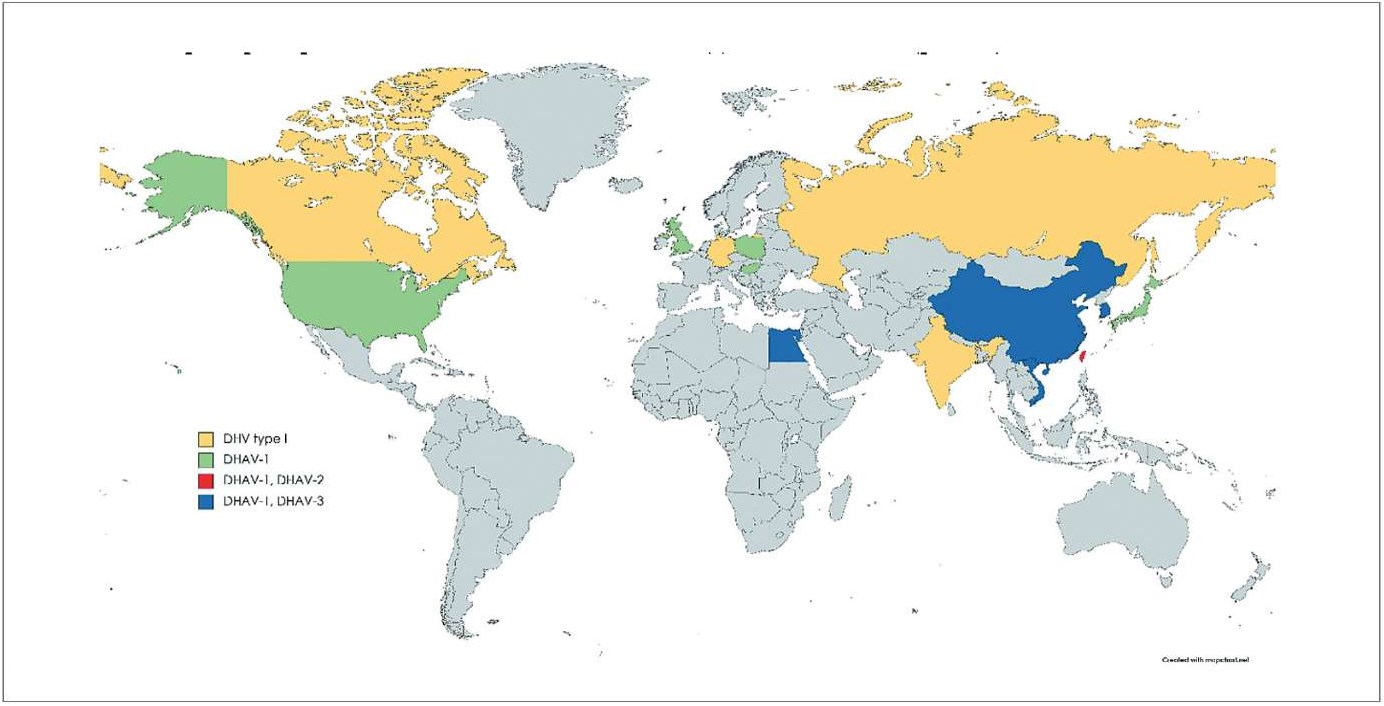
Рис. 1. Географическое распределение серотипов ВГУ (на 2021 г.) [10]
Fig. 1. Geographic distribution of DHV serotypes (for 2021) [10]
В настоящее время вспышки ВГУ продолжают регистрироваться в различных регионах мира, преимущественно в Азии, где заболевание наносит значительный экономический ущерб хозяйствам. Для Российской Федерации проблема также остается актуальной: учитывая наличие промышленного утководства, сохраняется риск заноса и распространения новых вариантов вируса.
Будучи представителем семейства пикорнавирусов, возбудитель ВГУ представляет собой небольшой безоболочечный вирус сферической формы размером 20–40 нм с одноцепочечной положительной РНК длиной около 7800 нуклеотидов. Полная открытая рамка считывания (ОРС), окруженная 5’ и 3’ некодирующими участками, кодирует три зрелых структурных белка, а именно белки капсида 0 (VP0), 1 (VP1) и 3 (VP3). ОРС также кодирует девять неструктурных белков (2A1–2A2–2A3–2B–2C–3A–3B–3C–3D), при этом основной поверхностный белок VP1 является антигенной детерминантой, играющей существенную роль в патогенности, эволюции и вирулентности вируса [6][10].
Исходя из особенностей генетической структуры возбудителя, он был разделен на три серотипа: серотип 1 (DHAV-A), который встречается повсеместно; серотип 2 (DHAV-B), выделенный на Тайване; и серотип 3 (DHAV-C), выделенный в Южной Корее и Китае. DHAV-C в основном поражает утят до 3-недельного возраста, его эпидемиология, клинические и патолого-анатомические признаки очень схожи с таковыми у DHAV-A, что затрудняет дифференциацию этих двух серотипов. При сравнении полных нуклеотидных и аминокислотных последовательностей VP1, VP0 и VP3, а также частичной трехмерной нуклеотидной последовательности были установлены различия между тремя серотипами вируса ВГУ [5].
Возбудитель ВГУ не проявляет гемагглютинирующей активности, культивируется на эмбрионах птиц (утиных, куриных, перепелиных, цесариных и гусиных), а также в первично трипсинизированных культурах клеток (фибробластов) указанных эмбрионов. Существуют данные об использовании первичных и перевиваемых культур клеток и различных опухолевых тканей. В культуре клеток вирус оказывает цитопатическое действие [7][14][15]. Выявлено, что тканевые культуры, происходящие от млекопитающих, и другие клетки, такие как НЕр-2, Детройт-6, HeLa, Л 99, ПМС, проявляют низкую чувствительность к вирусу ВГУ [14]. Установлено, что возбудитель ВГУ типа 2 хорошо реплицируется в утиных эмбрионах и организме уток, а также в культуре клеток, полученной из печени и почек утенка. При этом в отличие от вируса типа 1 его репродукция в культуре почек цыпленка и перепелки ограничена. Кроме этого, вирус типа 2 не реплицируется в эмбрионах кур. Утята, имеющие антитела к вирусу гепатита типа 1, заболевают при заражении вирусом гепатита типа 3. Особенности репродукции свидетельствуют о наличии антигенных различий у разных типов ВГУ [15].
Возбудитель ВГУ демонстрирует высокую степень устойчивости к воздействию внешней среды: в кормушках возбудитель может сохраняться более 10 нед., в подстилке – до 37 сут, в воде – до 74 сут, а в почве – от 105 до 157 сут, на поверхности стен птичников – от 20 до 40 сут, в зависимости от температуры воздуха. Весной вирус может оставаться жизнеспособным до 25 сут, а зимой – до 105 сут. Максимальный срок сохранения возбудителя в инфицированных водоемах составлял 74 сут [15].
Вирус устойчив к эфиру и хлороформу, а также к различным значениям pH среды. При хранении в холодильнике при температуре минус 14–32 °С возбудитель сохранял жизнеспособность в течение нескольких лет [15].
Высокие температуры оказывают губительное действие на вирус: нагревание до 56 °С вызывает его инактивацию в течение 60 мин. Ультрафиолетовые лучи уничтожают возбудитель в течение 10 мин [7].
Дезинфицирующие растворы ксилонафта, лизола, креолина и кальцинированной соды в обычных концентрациях неэффективны [15]. В птицеводческих хозяйствах для обеззараживания помещений применяют различные дезинфицирующие средства: 1%-й раствор формальдегида; 4%-й подогретый (40–45 °С) раствор гидроксида натрия, который выдерживают 12 ч; раствор гипохлорита натрия с содержанием 1,5% активного хлора и 1,2% свободной щелочи с экспозицией в 6 ч; кроме того, используют 5%-й раствор однохлористого йода, который выдерживают 6 ч [7].
Также для нейтрализации вируса можно использовать аэрозольную дезинфекцию птичников 40%-м водным раствором формальдегида при экспозиции 12 ч или 20%-м раствором при экспозиции 24 ч. Зараженный вирусом гепатита комбикорм можно обезвреживать посредством его запаривания при температуре 65–75 °С в течение 1 ч [7].
Эволюционные механизмы вируса гепатита утят схожи с механизмами других пикорнавирусов; основные общие черты этих вирусов включают высокую скорость эволюции и потенциал рекомбинации вирусной РНК даже между гетеротипичными штаммами. История эволюции и подтверждающие эпизоотологические данные позволяют предположить, что Азия может выступать в качестве очага диверсификации возбудителя ВГУ. Штаммы вируса ВГУ серотипа 1, циркулирующие в Европе, тесно связаны со штаммами серотипа 1, зарегистрированными в Китае. Данное заключение согласуется с предыдущими исследованиями других близкородственных патогенов, происходящих от водоплавающих птиц, географическое распространение которых совпадает между этими удаленными географическими районами. Все эти данные могут свидетельствовать о наличии общих факторов или способов распространения различных вирусов водоплавающих птиц между Дальним Востоком и европейскими регионами [10].
Заболевание поражает утят и гусят, в то время как птицы из отряда куриных и лабораторные животные к нему невосприимчивы [7]. Источником возбудителя инфекции является больная и переболевшая птица. Утята-реконвалесценты выделяют вирус с экскрементами, носовыми и конъюнктивальными секретами. Вирусоносительство после болезни может продолжаться от 60–75 до 300–650 сут. В естественных условиях заражение молодняка происходит при поедании инфицированного корма и воды, не менее важен и аэрогенный путь заражения [2][16]. Некоторые исследователи отмечают возможный вертикальный путь передачи возбудителя [17].
Характерной особенностью эпизоотий ВГУ, повторяющейся практически во всех случаях, является довольно быстрое нарастание гибели утят: пик приходится на 4–5-е сут, спад – на 7–8-е сут, к 10–12-м сут наблюдается резкое снижение числа погибших особей [15].
На течение заболевания влияют скученность молодняка, сырая подстилка, сквозняки, низкая температура помещения. Возможно заражение на водных выгулах при контакте домашней птицы с дикой [7].
Широкое распространение болезни обусловлено высокой устойчивостью вируса к физико-химическим факторам, его длительным присутствием в организме переболевшей птицы, генетической вариабельностью и устойчивой неблагополучной эпизоотической ситуацией по ВГУ в хозяйствах [16].
Вирусный гепатит утят за последние 6 лет регистрировали в Китае (2009–2021 гг.), Египте (2019 г.), Индии (2020–2023 гг.), Гайане (2019–2024 гг.) [18][19][20][21].
Появление новых генотипов и субгенотипов возбудителя ВГУ снижает эффективность применяемых вакцин. Из-за генетической изменчивости вируса многие существующие вакцины обеспечивают защиту только против определенных штаммов возбудителя, что делает их менее эффективными [22].
При заражении вирус проникает в организм через слизистые оболочки органов дыхания, желудочно-кишечного тракта и поврежденные участки кожи. Гепатоциты являются клетками-мишенями для возбудителя ВГУ – репродукция вируса происходит в цитоплазме печеночных клеток. Возбудитель накапливается не только в печени, но и в селезенке, фабрициевой сумке и головном мозге. У больной птицы развивается гепатит, отмечают некроз гепатоцитов, застойную гиперемию и кровоизлияния. Повреждение желчных протоков ведет к задержке эвакуации желчи. Происходит интоксикация организма. Затем развивается дистрофия сердечной мышцы, перикардит и нефрит, что приводит к гибели птицы [16]. Накопление вируса в селезенке и фабрициевой сумке указывает на то, что возбудитель может проникать как в центральные, так и в периферические иммунные органы. Это, в свою очередь, приводит к дисфункции иммунной системы у зараженных утят. Следовательно, даже после выздоровления от инфекции, вызванной вирусом гепатита, утята все равно остаются восприимчивыми к другим патогенам из-за ослабленной иммунной системы [3].
У инфицированных утят происходят изменения в обмене веществ: уменьшается концентрация общего белка, билирубина и альбумина, снижаются защитные свойства сывороточных коллоидов, щелочной фосфатазы, глутамат-пируват-трансаминазы, билирубина и креатинина, а также повышается активность ферментов сыворотки крови, что указывает на прямую связь с наличием повреждений печени у инфицированной птицы. Наблюдается некроз ацинарных клеток поджелудочной железы, что свидетельствует о влиянии на экзокринную функцию поджелудочной железы [3][15].
Инкубационный период может составлять 1–5 сут, он сокращается при пероральном, интраназальном и аэрогенном заражении по сравнению с парентеральным методом введения. Болезнь чаще всего протекает в сверхострой и острой форме. У заболевшей птицы наблюдают угнетение, отсутствие аппетита, нарушение координации движения, парезы, параличи, опистотонус (рис. 2), сужение глазной щели и конъюнктивит. Полное выздоровление отмечается редко. Переболевшие утята остаются вирусоносителями. При хроническом течении болезни птицы отстают в росте и развитии. Такое течение болезни чаще наблюдается у 3–4-недельных утят. Заболевание длится 10–20 сут, иногда более, сопровождаясь диареей. Утята становятся малоподвижными, у некоторых опухают суставы. Отмечается пингвиноподобная походка – птица передвигается, сохраняя вертикальное положение тела [15].

Рис. 2. Опистотонус при ВГУ (авторы фото: С. В. Глейзер, В. Ю. Фоменко, В. Н. Ирза)
Fig. 2. DНV-caused opisthotonus
(photo by: S. V. Glazer, V. Yu. Fomenko, V. N. Irza)
Патолого-анатомические признаки при хроническом течении болезни характеризуются увеличением печени в 1,5 раза, ее неравномерным окрашиванием, обнаруживаются гранулемы, характерные для лейкоза; селезенка кровенаполнена, с очагами некроза; развиваются периартриты [6][15].
Выжившие утята после выздоровления практически не отличаются от здоровых птиц. Однако из образцов патологического материала (печень, головной мозг), полученных от переболевших особей, можно изолировать вирус, а в образцах сыворотки крови выявить специфические антитела [6].
У взрослых уток заболевание протекает бессимптомно, иногда отмечается овариосальпингит [7].
Заболеваемость среди утят в возрасте до трех недель составляет 80–90%. При сверхостром течении болезни летальность до 10-суточного возраста может достигать 100%, а при остром течении – 70–80%. В стационарно неблагополучных хозяйствах вирусный гепатит регистрируется среди утят от 15–30-суточного возраста и старше. Падеж в отдельных партиях может составлять 5–10%. При повторном завозе утят, не имеющих антител к ВГУ, в такое хозяйство летальность от партии к партии увеличивается и достигает иногда 80–95% [8]. Эмбрионы, пораженные вирусом ВГУ, в 75–90% случаев погибают на различных стадиях эмбрионального развития [16].
Вирусный гепатит утят нередко осложняется одновременным развитием других заболеваний вирусной, бактериальной и грибковой этиологии, таких как грипп, сальмонеллез, микоплазмоз, колибактериоз, аспергиллез. Важно отметить, что в развитии и протекании такой ассоциированной инфекции ВГУ играет ключевую роль [15].
При вскрытии отмечают поражение печени (рис. 3): она имеет дряблую консистенцию, увеличена, неравномерно окрашена в охряно-желтый или серовато-глинистый цвет, с большим количеством кровоизлияний под капсулой и в паренхиме. Желчный пузырь переполнен желчью. Селезенка иногда увеличена и имеет крапчатый вид. В большинстве случаев почки увеличены, кровенаполнены, почечные кровеносные сосуды гиперемированы. Сосуды головного мозга полнокровны. Сердце имеет вид вареного мяса, в состоянии гранулярной дистрофии, коронарные сосуды наполнены кровью. Легкие отечны, наблюдаются фибринозно-дифтеритические наложения на стенке воздухоносного мешка [7][16]. Часто у молодняка наблюдается катаральное воспаление слизистой оболочки кишечника, что является следствием гепатита, осложненного бактериозом, в первую очередь сальмонеллезом [15].
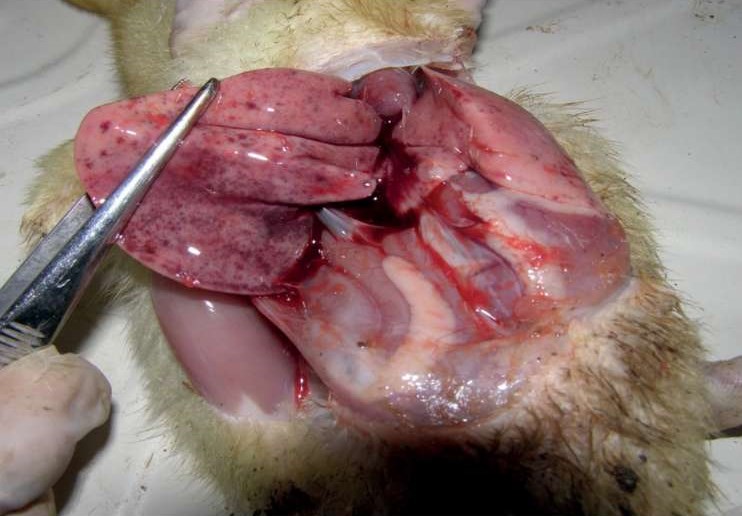
Рис. 3. Поражение печени при ВГУ
(авторы фото: С. В. Глейзер, В. Ю. Фоменко, В. Н. Ирза)
Fig. 3. DHV-caused liver lesions
(photo by: S. V. Glazer, V. Yu. Fomenko, V. N. Irza)
При гистологическом исследовании пораженной печени наблюдают некроз гепатоцитов с полным кариолизисом, зернистой дистрофией, жировым метаморфозом и лимфоцитарно-плазмоцитарной пролиферацией, реже – гиперемией кровеносных сосудов межбалочных капилляров. При окраске гистосрезов печеночной ткани по методу Нобля татразинфлоксином можно обнаружить тельца-включения овальной или шаровидной формы размером от 1 до 8 мкм [7][16]. В гепатоцитах уже в первые 1–18 ч после заражения можно обнаружить вирусоподобные частицы. По истечении суток в печени начинают наблюдаться первые признаки изменения в виде некробиоза и апоптоза гепатоцитов, в ряде случаев отмечают обширные некротические очаги, геморрагии. Впоследствии внутри печеночных долек и вблизи синусоидных капилляров выявляются мелкоочаговые пролифераты и распад пораженных клеток. Вокруг кровеносных сосудов и желчных протоков наблюдается периваскулярная инфильтрация, которая представлена гранулоцитами, лимфоцитами и плазматическими клетками. В период выздоровления птицы в паренхиме печени можно обнаружить начало регенеративных процессов [6].
В селезенке наблюдают активные регрессивные изменения, переходящие в некротические [7][16]. В фабрициевой сумке после заражения отмечаются определенные патоморфологические изменения: гиперемия, повышенная инфильтрация серозным экссудатом, пролиферация плазматических клеток и гемоцитобластов. Пролиферация становится выраженной спустя 72 ч после инфицирования, что связано с иммунологической реакцией на антиген вируса. Если инфекция длится 2–3 нед., то количество плазматических клеток в фабрициевой сумке и селезенке увеличивается в 3–4 раза. В тимусе отсутствуют какие-либо существенные патолого-анатомические изменения. Характер изменений в головном мозге указывает на серозный энцефалит [6].
У эмбрионов, зараженных вирусом гепатита и замерших на 20–25-е сут инкубации, регистрируют кровенаполнение сосудов желточного мешка, застойные явления и отечность подкожной клетчатки в области головы, шеи и спины. В отдельных случаях отечность наблюдается в области конечностей, часто встречаются множественные кровоизлияния. У эмбрионов, погибших через 15 сут после начала инкубации, регистрируют гиперемию сосудов желточного мешка и аллантоиса. В аллантоисной полости присутствует прозрачная тягучая, иногда опалесцирующая жидкость зеленоватого цвета. Часто у эмбрионов наблюдается явление «карликовости», а в некоторых случаях – атрофия мышц конечностей. Печень павших эмбрионов увеличена, имеет неравномерный цвет, который варьирует от светло-коричневого до темно-зеленого цвета. Из-за чередования пораженных и нормальных участков паренхимы печень приобретает мраморный вид. На поверхности печени можно обнаружить очаги некроза [6].
Диагноз на ВГУ ставят на основе эпизоотологических, клинических, патолого-анатомических данных и результатов лабораторных исследований.
При патолого-анатомическом исследовании павших утят основное внимание при остром течении болезни уделяют наличию геморрагического и некротического гепатита, гломерулонефрита, серозного энцефалита и миокардиодистрофии. При подостром течении типичным признаком гепатита утят является катаральный энтерит. У утят до 3-недельного возраста патогномоничным признаком является наличие геморрагий различной формы и интенсивности на всей поверхности печени [6].
При проведении лабораторных исследований сначала выделяют изолят вируса на чувствительных биологических системах. Для этого из исследуемых проб печени, селезенки и головного мозга 10–15 трупов утят готовят 10–20%-ю суспензию. Полученный надосадок инъецируют утятам 2–14-суточного возраста или инокулируют утиным и куриным эмбрионам 9–12- и 8–10-суточного возраста соответственно, или вносят в первичную культуру клеток почек, печени и фибробластов утиных и куриных эмбрионов. У погибших эмбрионов аллантоисная жидкость и содержимое желточного мешка приобретают зеленоватый оттенок. На поверхности тела эмбриона отмечают кровоизлияния, а также отечность грудной и брюшной полостей с наличием кровоизлияний. Печень имеет неплотную, рыхлую консистенцию и серо-желтый цвет, наблюдаются очаги некротических изменений. Также у павших эмбрионов отмечают нефрозо-нефрит [15]. Утиные эмбрионы демонстрируют бóльшую восприимчивость, чем куриные, которая характеризуется более ранним замиранием развития и более выраженными изменениями. При проведении повторных пассажей выраженность перечисленных поражений усиливается. Эмбрионы, замершие на 5–8-е сутки после заражения, часто характеризуются уменьшенными размерами – признаком «карликовости» [6].
Для быстрой диагностики ВГУ типа 1 можно использовать непрямой метод флуоресцирующих антител (МФА). Для идентификации выделенного вируса – реакцию нейтрализации (РН) на утятах, эмбрионах и культурах клеток. Для ретроспективной диагностики ряд авторов применяли реакцию непрямой гемагглютинации (РНГА), непрямой иммуноферментный анализ (ИФА), реакцию диффузионной преципитации (РДП), реакцию встречного иммуноэлектроосмофореза (РВИЭОФ) [16].
В 1982 г. впервые была описана реакция ингибирования бляшек вируснейтрализующими антителами. Данный метод продемонстрировал гораздо более высокую чувствительность, чем РН вируса в эмбрионах.
Также в 1988 г. была описана реакция микронейтрализации в культуре клеток почек утиного эмбриона для серологической диагностики ВГУ типа 1. В то время данный метод был практичнее, быстрее и экономичнее по сравнению с другими альтернативными методами, но метод ингибирования бляшек являлся более чувствительным.
В лабораторной диагностике эти методы популярны благодаря простоте их исполнения и доступности. Однако следует учитывать, что они имеют ряд недостатков. Если оценивать их по таким параметрам, как чувствительность, специфичность и быстрота получения окончательного результата, то вышеупомянутые способы диагностики не являются оптимальными.
В настоящее время для определения антител к возбудителю ВГУ типа 1 чаще всего применяют метод ИФА. При оценке различных методов выявления антител к вирусу гепатита утят типа 1 в сыворотке крови было установлено, что твердофазный ИФА по степени чувствительности близок к РН, значительно менее чувствительной была РДП [6].
Разработанный в 1998 г. метод ИФА для выявления специфических антител к вирусу гепатита утят типа 1 в сыворотке крови оказался высокочувствительным и специфичным. В связи с ключевой ролью гуморального иммунитета при инфицировании DHАV-1 S. Mao et al. разработали непрямые методы ИФА для выявления сывороточных иммуноглобулинов: IgG, IgM и IgA. Результаты, полученные авторами, показали, что корреляция между непрямым ИФА и РН составила 95,2% для IgG и IgA, и 75% для IgM. Согласно мнению авторов, новый непрямой ИФА представляет собой перспективный и практичный метод для оценки гуморального ответа на DHAV-1. Данный метод может быть полезен в исследованиях, направленных на изучение иммунного ответа при заражении вирусом гепатита [23].
В настоящее время предложены косвенные варианты ИФA, в которых белки VP1 и VP3 DHAV-1 применяют в качестве сорбирующих антигенов для выявления специфических антител к вирусу. Данный вариант ИФА быстрее, проще и практичнее, чем классический. Была разработана иммуноферментная тест-система (VP1-ELISA), предназначенная для выявления антител к вирусу гепатита утят типа 1. В качестве сорбирующего антигена применялся рекомбинантный белок VP1, клонированный и экспрессированный в Escherichia coli. Данный метод продемонстрировал высокую специфичность (92,5%) и чувствительность (96,7%) по сравнению с РН. Благодаря специфичности белка VP1 для распознавания антител к DHAV-1, VP1-ELISA не реагирует с антисывороткой на другие вирусные инфекции уток. Разработчики полагают, что предложенная VP1-ELISA представляет собой высокочувствительный и специфический тест, пригодный для скрининга на наличие инфекции, вызываемой DHАV-1, а также для мониторинга уровня антител к вирусу гепатита утят типа 1 [24].
Впервые непрямой ИФА с использованием рекомбинантного белка VP3 вируса гепатита утят типа 1 (DHАV-1) создали Y. Shen et al., новый метод позволяет обнаруживать антитела сразу к DHAV-1 и DHAV-3. Данный метод на основе VP3 характеризуется высокой чувствительностью, специфичностью и воспроизводимостью и по эффективности не уступает методу непрямого ИФА на основе вируса гепатита утят типа 1 для серологических исследований [25].
Прогресс в области иммунологии и молекулярной биологии привел к появлению инновационных лабораторных методов диагностики. Одним из таких методов является полимеразная цепная реакция (ПЦР), которая в настоящее время получила широкое распространение для обнаружения и определения DHAV-1 [6].
В Китае ВГУ ассоциируется с тремя разновидностями вирусных агентов: DHAV-1, DHAV-3 и DAstV-1 [26][27][28]. Независимо от того, каким из этих типов вируса заражены утята, у них проявляются схожие признаки: короткий инкубационный период, стремительное развитие болезни, высокая летальность, опистотонус и увеличение печени с обильными кровоизлияниями. Предварительный диагноз «ВГУ типа 1» ставят на основании клинических признаков и результатов патолого-анатомического вскрытия, однако установить, какой тип вируса или их комбинация вызвала инфекцию, затруднительно. В 2008 г. для различия штаммов DHAV-1 и DHAV-3 была разработана мультиплексная ПЦР [29]. Относительно недавно ученые усовершенствовали метод, разработав дуплексный тест: ПЦР с обратной транскрипцией (ОТ-ПЦР, RT-PCR), что упрощает оперативное и экономичное выявление в лаборатории смешанных инфекций, вызванных различными штаммами DHAV-1 и DHAV-3 у утят [30]. Однако ни один из указанных выше ПЦР-анализов не позволял обнаружить DAstV-1.
Chen L. et al. впервые разработали мультиплексную ОТ-ПЦР, способную одновременно идентифицировать DHAV-1, DHAV-3 и DAstV-1 в клинических образцах. Результаты показали, что данный метод обладает высокой специфичностью для вируса ВГУ и не обнаруживает генетический материал других утиных патогенов. Важно отметить, что дифференциальная диагностика DHAV-1, DHAV-3 и DAstV-1 может быть выполнена в течение нескольких часов в одной реакции в отличие от классических методов, включающих тесты с перекрестной нейтрализацией, которые занимают несколько дней. Таким образом, данный мультиплексный ПЦР-анализ представляет собой быстрый, эффективный и практичный метод для дифференциальной диагностики смешанных инфекций, вызванных тремя типами вирусов ВГУ, а также для эпизоотологического мониторинга ВГУ типа 1 [31].
Современная молекулярно-биологическая диагностика ВГУ типа 1 постоянно развивается и обогащается новыми методиками. Так, учитывая изменчивость эпизоотической ситуации в Юго-Восточной Азии, где циркулирует 2 серотипа вируса гепатита утят (DHАV-1 и DHАV-3), был разработан дуплексный анализ ПЦР в режиме реального времени (ОТ-ПЦР-РВ; qRT-PCR), позволяющий одновременно и количественно определять наличие в образцах DHAV-1 и DHAV-3 [32].
Также был разработан мультиплексный метод ПЦР для быстрого обнаружения нескольких вирусных патогенов уток: вирус гепатита утят А типа 1, вирус чумы утки, парвовирус и реовирус мускусной утки и вирус гриппа птиц подтипа H9N2. Результаты показали, что мультиплексная ПЦР-система эффективна для выявления вирусных нуклеиновых кислот в эмбрионах уток, инфицированных шестью распространенными вирусами, а также в клинических образцах. Исследователи утверждают, что созданный ими мультиплексный метод ПЦР обеспечивает специфическое, чувствительное и высокопроизводительное обнаружение указанных шести утиных вирусов и может быть использован для клинической идентификации и диагностики вирусной инфекции уток [33].
Необходимо подчеркнуть, что имеющиеся молекулярно-биологические методы не позволяют дифференцировать вакцинный штамм вируса DHАV-1 от полевого. Только K. P. Li et al. создали метод ОТ-ПЦР-РВ и анализ кривых плавления с высокой разрешающей способностью (HRM) для оперативного выявления и дифференциации вакцинного и полевого штаммов DHАV-1. Разработанный метод демонстрирует высокую специфичность к DHAV-1 и способен обнаруживать около 100 копий вирусной РНК. Исследователи продемонстрировали возможность обнаружения вируса гепатита А типа 1 в образцах фекалий утят уже через 6 ч после инфицирования. На основании этого авторы делают вывод о потенциальной пользе разработанного ОТ-ПЦР-РВ и HRM для диагностики и мониторинга распространения инфекции, обусловленной DHAV-1 [34].
Иммунный ответ при ВГУ включает врожденные, гуморальные, клеточные и материнские механизмы защиты.
Врожденный иммунный ответ служит первой линией защиты и инициируется быстрым распознаванием вирусных компонентов паттерн-распознающими рецепторами (PRR), включая толл-подобные рецепторы 7 и 3 (TLR7, TLR3), индуцируемый ретиноевой кислотой ген 1 (RIG-1) и ген 5, ассоциированный с дифференцировкой меланомы (MDA5), которые обычно участвуют в борьбе с инфекцией РНК-вирусов. У утят, инфицированных вирусом гепатита, выявляется значительная индукция интерферонов типов I и II, а также провоспалительных цитокинов (IL-2, IL-6), причем экспрессия некоторых из них зависит от возраста птицы. Таким образом, врожденный иммунитет определяет исход ранних стадий инфекции и влияет на последующее развитие иммунного ответа [35].
Клеточный ответ формируется после представления антигена на поверхности макрофагов и дендритных клеток, которые активируют Т-лимфоциты. Процесс клеточного иммунитета связан с активацией множества компонентов: специфических рецепторов лимфоцитов, главного комплекса гистосовместимости, дендритных лимфофолликул, Т- и В-клеток, переходящих из фазы G0 в G1, а также цитокинов и лимфокинов, обеспечивающих передачу сигналов, что в итоге приводит к элиминации антигена [8]. Т-лимфоциты играют ключевую роль в уничтожении инфицированных клеток печени и формировании иммунологической памяти. Сильный ответ CD8+ Т-клеток, продуцирующих IFN-γ, ассоциирован с элиминацией вируса, тогда как утята с подавленной функцией Т-клеток переносят инфекцию тяжелее, что подтверждает значение клеточного звена защиты [35].
Ключевым фактором защиты является выработка вируснейтрализующих антител, преимущественно направленных против капсидного белка VP1, который является основным иммуногенным компонентом возбудителя ВГУ [36]. В сыворотке крови утят нейтрализующие антитела появляются уже на 4-е сут после иммунизации, а максимальный уровень формируется через 7–9 сут [8]. Однако исследования динамики специфических антител к вирусу гепатита остаются ограниченными, в основном концентрируясь на разработке методов их детекции и оценке степени защиты после вакцинации [35].
Существенную роль играет пассивная защита, обес-печиваемая потомству через желток яиц от вакцинированных или переболевших уток. У суточных утят иммунная система еще недостаточно зрелая и потому не способна полностью противостоять инфекционным болезням. Приобретенный иммунитет формируется постепенно в течение первых нескольких недель после вылупления, поэтому передача материнских антител играет важную роль в защите потомства. Антитела из желточного мешка поступают в кровь развивающегося эмбриона и сохраняются у вылупившегося утенка. Данный процесс лежит в основе стратегий вакцинации птиц в промышленном птицеводстве для борьбы с инфекционными заболеваниями. Вакцинация молодняка до начала яйцекладки позволяет сформировать достаточный уровень защитных антител, который сохраняется в течение всего периода их продуктивности и, как следствие, обеспечивает передачу материнских антител потомству [6]. Пассивно полученные антитела сохраняются у утят в течение 2–3 нед., формируя «иммунологическое окно», требующее ранней вакцинации. В экспериментах показано, что через 5 нед. после иммунизации уток родительского стада титры антител достигали (8,4 ± 1,3) log2 и сохранялись на уровне (9,0 ± 1,1) log2 до 36 нед., при этом передача антител потомству составила (12,8 ± 3,0)%, а их период полувыведения у утят был около (3,4 ± 1,1) сут [37]. Утята, полученные от иммунных уток, а также молодняк старше 6-недельного возраста, как правило, невосприимчивы к вирусу в 70–100% случаев [38].
Для лечения можно использовать сыворотку и плазму крови реконвалесцентов, которую готовят после убоя взрослой птицы. Ранее использовали гипериммунную сыворотку крови лошадей и взрослых уток, но из-за низкой эффективности данного метода от него отказались [7][39].
В качестве неспецифической профилактики требуется проводить необходимые ветеринарно-санитарные мероприятия, обеспечивать изолированное содержание молодняка, осуществлять регулярную дезинфекцию яиц, инкубаторов, помещений и водоемов, ограничивать контакт с дикими птицами [7]. Однако важнейшей мерой профилактики является вакцинация птицы для создания напряженного иммунитета.
Основные характеристики существующих вакцин против вирусного гепатита утят представлены в разделе Дополнительные файлы по адресу https://doi.org/10.29326/2304-196X-2026-15-2-110-122.
Наиболее широко применяются живые аттенуированные вакцины для иммунизации утят в возрасте 1–3 сут. Они обеспечивают продолжительный иммунитет и защиту с первых дней жизни, что особенно важно, учитывая возрастную восприимчивость к возбудителю ВГУ. Аттенуацию выделенных изолятов вируса проводят путем многократных пассажей в различных биологических системах (куриных и утиных эмбрионах, первичных культурах клеток фибробластов куриных и утиных эмбрионов и др.) [39].
В Российской Федерации живую эмбриональную вакцину против ВГУ из штамма «ВГНКИ-К» производит ФГБУ «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ»). Через 48 ч после вакцинации у птицы формируется иммунитет, который у утят сохраняется не менее 30 сут после однократного применения, у уток – не менее 6 мес. после двукратного применения [40]. Во Вьетнаме выпускают вакцину AVAC DVH Live, изготовленную из аттенуированного штамма DHAV-1, культивированного на куриных эмбрионах, с активностью не менее 10³,³ ЭЛД50 в одной дозе. Она обеспечивает иммунитет уже на 3–5-е сут и полную защиту к 7-м сут после вакцинации [41]. Против ВГУ генотипа 1 за рубежом производят модифицированную живую вакцину (США), Hepatovax (Merial, Франция) и живую вакцину из штамма CH60 (Huapai Biotechnology Group, Китай). Вирус в составе вакцины, выпускаемой в США, был ослаблен путем последовательного пассирования на куриных эмбрионах. Эта вакцина вводится однократно в дозе 0,5 мл под кожу в области шеи суточным утятам, не имеющим материнских антител против вируса гепатита. Однако продолжительность иммунитета у утят не определена. Кроме того, вакцину можно вводить уткам-несушкам в дозе 0,5 мл под кожу в области шеи на 16, 20 и 24-й нед., а затем каждые 12 нед. в течение всего периода яйцекладки. Три первые иммунизации необходимы для пассивной защиты утят, которая может сохраняться в течение 2–4 нед. после вылупления. Живая вакцина из штамма CH60 была получена после 60 последовательных пассажей на куриных эмбрионах. Одна доза (0,25 мл) этой вакцины вводится утятам в возрасте от 1 до 7 сут внутримышечно в область бедра, а продолжительность иммунитета составляет более 1 мес. Уток-несушек можно вакцинировать за 1 нед. до начала яйцекладки одной дозой (0,25 мл), что обеспечивает пассивный иммунитет у потомства на 6 мес. [42].
Кроме того, против ВГУ генотипа 3 были разработаны живые вакцины из штамма AP-04203-P100 (Республика Корея), прошедшего 100 пассажей на куриных эмбрионах, и из штамма SD70 (Китай), прошедшего 70 пассажей на куриных эмбрионах, с титром вируса 10⁷,⁵ ЭЛД50/мл. Вакцина из штамма SD70 не вызывала клинических признаков у утят суточного возраста и защищала от заражения DHAV-3 при однократной инъекции [43][44].
В ряде стран, где одновременно циркулируют генотипы 1 и 2 ВГУ, была разработана бивалентная живая аттенуированная вакцина DHAV-1 + DHAV-3, обеспечивающая защиту утят от обоих генотипов вируса. В суточном возрасте птицам вводили двухвалентную вакцину посредством внутримышечной инъекции. Иммунизированные утята были устойчивы к заражению вирулентными штаммами DHAV-1 и DHAV-3 через двое или трое суток после вакцинации. Более того, у утят регистрировали стойкий гуморальный иммунный ответ, который достигал пика через 3 нед. и сохранялся в течение 6 нед. после вакцинации [45].
Изучалась эффективность перорального введения живой вакцины в качестве бустерной иммунизации после первичного внутримышечного введения инактивированной вакцины. У племенных уток вируснейтрализующие антитела вырабатывались в высоких титрах и сохранялись в течение 36 нед., ни одно из потомств не погибло от заражения вирусом в возрасте 1, 7 и 14 сут. Эти результаты свидетельствуют о том, что живые вакцины против ВГУ можно использовать для выработки высокого уровня антител и формирования стойкого иммунитета у производителей, а также обеспечения надежного пассивного иммунитета у потомства [46]. Преимущество данного метода состоит в возможности массового введения вакцины с питьевой водой, что повышает практичность и эффективность иммунизации в хозяйствах. Тем не менее живые вакцины против ВГУ в основном инъецируют внутримышечно или подкожно, тогда как исследования по пероральному способу введения пока ограничены.
Молекулярные механизмы аттенуации вируса изучались при проведении серийных пассажей. Анализ геномной последовательности штамма DHAV-1 после 40 пассажей в куриных эмбрионах показал постепенное накопление нуклеотидных и аминокислотных мутаций. Большая часть этих изменений наблюдалась в белках 2C, 3D и VP1. Кроме того, 100 последовательных пассажей штамма DHAV-3 в куриных эмбрионах привело к стабильному накоплению мутаций в различных участках, включая 3D, VP2, 2C и 3’-нетранслируемую область. В аттенуированном штамме DHAV-3, полученном после 70 пассажей в куриных эмбрионах, было выявлено в общей сложности 12 аминокислотных замен и три нуклеотидные мутации. Примечательно, что мутация Y180C в белке VP1 и мутация S286N в белке 2C привели к утрате потенциальных участков гликозилирования. Сравнение генов VP1 полевого и аттенуированного штаммов DHAV-1 выявило ряд устойчивых аминокислотных замен, которые, возможно, связаны с ослаблением вируса. Следует отметить, что все эти мутации были обнаружены путем секвенирования аттенуированных штаммов возбудителя ВГУ после серии пассажей, их непосредственная роль в ослаблении вируса не подтверждена. Детальное понимание молекулярных основ вирулентности возбудителя ВГУ может стать основой для разработки новых живых аттенуированных вакцин [42][44][47].
Еще одной потенциальной проблемой живых вакцин является безопасность. В связи с тем что производство живых вакцин против ВГУ предполагает многократное пассирование вирулентных штаммов в эмбрионах кур или уток, необходимо тщательно оценивать безопасность этих вакцин. Несмотря на то что некоторые исследования демонстрировали отсутствие возврата вирулентности после нескольких циклов обратных пассирований живой вакцины DHAV-3 на утках, в других работах показано сохранение вирулентности штамма DHAV-3 даже после 80 пассажей на куриных эмбрионах, утрата патогенности наблюдалась лишь после 90 пассажей. Важно отметить наличие задокументированных случаев восстановления вирулентности аттенуированного вируса гепатита после пассирования на утках. Аналогичная ситуация наблюдалась с двумя штаммами возбудителя ВГУ, которые, несмотря на 90 пассажей в куриных эмбрионах, восстановили свою вирулентность при последовательном пассировании на утятах. Следовательно, продолжительное пассирование вируса ВГУ в куриных эмбрионах может приводить к значительной аттенуации возбудителя, но вероятность возврата вирулентности, особенно в полевых условиях, исключать нельзя. В связи с этим необходим постоянный мониторинг безопасности живых вакцин против ВГУ [42][44].
Производство живых вакцин против ВГУ, как и многих других живых вакцин против болезней птиц, в значительной степени зависит от наличия свободных от патогенной микрофлоры (СПФ) эмбрионов. Учитывая некоторые недостатки такого производства, исследователи вели разработки по созданию стабильных клеточных линий, подходящих для размножения вируса ВГУ. В частности, установлено, что DHAV-1 способен к репликации и вызывает цитопатические изменения в модифицированной клеточной линии, полученной из фибробластов утиного эмбриона, что свидетельствует о ее пригодности к производству живых вакцин против ВГУ. Также была разработана линия клеток гусиного эмбрионального эпителия (GEE), в которой DHAV-1 демонстрирует способность к размножению в высоких титрах, что свидетельствует о перспективности использования GEE для производства живой вакцины [48]. Однако стоит отметить, что репликация вируса в этих клеточных линиях оценивалась в небольших объемах в лабораторных условиях. Данных о крупномасштабном производстве вакцин против ВГУ с использованием клеточных культур пока нет. Важно учитывать, что сыворотка крови плода теленка, используемая в культивировании клеток, может оказывать ингибирующее действие на репликацию DHAV-1 и DHAV-3, подавляя связывание вируса с клетками и его высвобождение. В связи с этим платформа бессывороточной клеточной культуры представляется перспективным направлением для будущего производства живых вакцин против ВГУ с использованием клеточных культур [42][49][50].
Живые вакцины против ВГУ имеют ряд преимуществ: они отличаются хорошей переносимостью для утят, быстрым формированием иммунитета, возможностью массового введения через питьевую воду и индукции гуморального и клеточного иммунитета. Живые вакцины также имеют ряд недостатков, таких как длительный процесс скрининга аттенуированных вакцинных штаммов, риск (даже очень низкий) реверсии вирулентности, потребность в соблюдении температурного режима хранения, высокая стоимость СПФ-эмбрионов. Следует отметить, что благодаря быстрому формированию иммунитета живые вакцины являются основным выбором для профилактики ВГУ у утят. Живые вакцины также могут использоваться в качестве бустерного компонента после первичной иммунизации племенного молодняка инактивированными вакцинами [42].
Инактивированные вакцины применяются в основном для иммунизации родительского поголовья с целью передачи материнских антител потомству. Отечественные производители инактивацию вируса ВГУ проводят аминоэтилэтиленимином или биоцидом «Инак», а в качестве адъюванта используют масляные эмульсии или гидроокись алюминия [51][52][53][54][55].
Ряд исследований свидетельствует о более высокой эффективности живых вакцин против ВГУ по сравнению с инактивированными. Было показано, что ни одна из инактивированных вакцин при использовании их в качестве первичной иммунизации у 16-недельных уток не вызывала выработки антител на достаточном уровне. Титры антител повышались только тогда, когда вакцина вводилась уткам трехкратно в возрасте 8, 16 и 22 нед., после этого они обеспечили защиту потомству в возрасте до 3 нед. Напротив, у уток, вакцинированных живой вакциной в возрасте 2–3 сут, вырабатывались стойкие антитела на высоком уровне, сохранявшиеся до 30 нед., а утята, выведенные от этих уток, были устойчивы к заражению вирусом до 3-недельного возраста. Также сообщалось, что иммуногенность инактивированной вакцины против ВГУ у племенных уток была низкой, но повышалась при первичной иммунизации живыми вакцинами [42]. Однако другое исследование показало, что бивалентная инактивированная вакцина против ВГУ типов 1 и 3 обладает высокой иммуногенностью и эффективностью, а иммунитет у утят сохраняется более 5 нед. [56]. Причины различий между результатами этих исследований неизвестны. На зарубежном рынке представлены как минимум две коммерческие инактивированные вакцины против гепатита утят, в том числе Orniduck (Bioveta, Чехия) и бивалентная вакцина против ВГУ генотипа 1 и 3 (Yebio, Китай) [42].
Различные группы исследователей для создания векторных вакцин работали над экспрессией генов протективных антигенов возбудителя ВГУ, таких как ген VP0 вируса гепатита серотипа 1, ген VP1 вируса серотипа 1 или 3, а также гены P1 и 3C вируса серотипа 3, в различных участках генома вируса энтерита уток. Рекомбинантные вирусы обладали высокой иммуногенностью против ВГУ и вирусного энтерита уток, что указывает на их потенциал для использования в качестве бивалентных вакцин для уток [57][58][59]. В некоторых исследованиях сообщалось, что рекомбинантные птичьи аденоассоциированные вирусы, экспрессирующие гены VP1 или VP3 возбудителя ВГУ серотипа 1, индуцировали защитный иммунитет у уток [60][61]. Более того, гены VP1 вируса ВГУ серотипов 1 и 3 коэкспрессировались в векторе вируса ньюкаслской болезни, и рекомбинантный вирус вызывал выработку вируснейтрализующих антител у уток [62]. Векторные вакцины обладают рядом преимуществ, включая стабильную экспрессию чужеродных антигенов, отсутствие необходимости в концентрировании антигена, формирование гуморального, клеточного или мукозального иммунитета, высокую генетическую стабильность и безопасность. Однако для получения таких вакцин требуется сложная молекулярная инженерия генома вируса и специальные технологии [42].
Рекомбинантные субъединичные белковые вакцины представляют собой тип вакцин, содержащих защитные компоненты патогенов, которые могут вызывать иммунный ответ у хозяина. В связи с недостатками системы производства вакцин, сырьем для изготовления которых служат яйца, субъединичные белковые вакцины являются потенциальной альтернативой традиционным вакцинам против ВГУ.
Белок VP1 вируса гепатита является основной мишенью нейтрализующих антител и участвует в связывании рецепторов, и поэтому этот протеин обычно экспрессируется как защитный антиген в различных системах [63]. Белок VP1 возбудителя ВГУ типа 1 был экспрессирован в Pichia pastoris, что индуцировало гуморальный и клеточный иммунный ответ у утят [64]. Кроме того, VP1 вируса гепатита серотипа 3 был экспрессирован в E. coli в качестве вакцинного антигена, но он обеспечивал лишь 25%-ю защиту от заражения вирусом. Смесь VP1 с флагеллином значительно усилила гуморальный, клеточный и цитокиновый ответ у утят и обеспечила 75%-ю защиту [65]. Преимущества рекомбинантных белковых субъединичных вакцин включают возможность крупномасштабного производства в клеточных системах, низкую стоимость и сопоставимую с цельновирусными вакцинами иммуногенность. Однако может потребоваться очистка антигена и систематическая оптимизация для повышения выхода антигена в различных системах экспрессии.
Другой тип вакцин против ВГУ – это вакцины на основе вирусоподобных частиц (ВПЧ). Они напоминают естественные вирусные частицы по морфологии и размеру, но не содержат нуклеиновых кислот. Такие вакцины безопасны для животных, поскольку ВПЧ не способны реплицироваться в организме хозяина, и индуцируют клеточный и гуморальный иммунитет. Исследования, посвященные вакцинам против ВГУ на основе ВПЧ, ограничены, но аналогичная стратегия используется для создания вакцин на основе ВПЧ для различных пикорнавирусов. Ген P1 кодирует структурные белки пикорнавируса, а полипротеин P1 расщепляется вирусной протеазой 3CD на капсидные белки, которые собираются, образуя пустые вирусные капсиды [66]. Таким образом, общая стратегия получения ВПЧ пикорнавирусов, таких как полиовирус, энтеровирус типа 71 и вирус Коксаки A16, заключается в совместной экспрессии генов P1 и 3CD в клетках насекомых или растениях [67][68][69]. Аналогичным образом A. Wang et al. показали, что ВПЧ вируса гепатита типа 1 могут образовываться в клетках насекомых, инфицированных рекомбинантным бакуловирусом, коэкспрессирующим гены P1 и 3CD. Иммунизация вакциной на основе ВПЧ вызывала устойчивый гуморальный ответ и обеспечивала надежную защиту [70]. Хотя такие вакцины являются многообещающей платформой для разработки новых вакцин против ВГУ, экспериментальные параметры должны быть оптимизированы для максимального выхода ВПЧ, а очистка антигена может привести к увеличению затрат.
Вирусный гепатит утят является значимой инфекцией, наносящей большой экономический ущерб утководческим хозяйствам. Для предупреждения возникновения ВГУ в хозяйствах необходимо проводить вакцинопрофилактику поголовья.
Несмотря на наличие широкого спектра зарубежных вакцин, на сегодняшний день в Российской Федерации единственным официально зарегистрированным препаратом для профилактики ВГУ остается живая вакцина из штамма «ВГНКИ-К», производимая ФГБУ «ВНИИЗЖ» [71]. Вакцина позволяет снижать уровень заболеваемости и смертности молодняка, но имеет ограничение в сроке хранения (9 мес.). В настоящее время в учреждении проводятся исследования по увеличению срока хранения препарата. Вакцину планируется выпускать в лиофилизированном виде, что обеспечит более высокую стабильность препарата.
Вклад авторов: Куникова Е. Д. – подготовка текста статьи, проведение поисково-аналитической работы; Волков М. С. – научное руководство, редактирование статьи, концепция обзора; Мороз Н. В. – редактирование статьи.
Contribution of the authors: Kunikova E. D. – text preparation, search and analysis; Volkov M. S. – scientific supervision, paper editing, review concept; Moroz N. V. – paper editing.
1. Трефилов Б. Б., Никитина Н. В., Явдошак Л. И., Трубицын М. М. Сравнительная оценка антигенности живой и инактивированной вакцины против вирусного гепатита утят типа I. Ветеринария. 2019; (4): 24–27. https://doi.org/10.30896/0042-4846.2019.22.4.24-27
2. Трефилов Б. Б., Никитина Н. В., Дмитриев К. Ю., Трубицын М. М. Вирусный гепатит утят типа I (эпизоотология, патогенез и диагностика). Эффективное животноводство. 2017; (3): 16–17. https://elibrary.ru/zivrzn
3. Niu Y., Ma H., Ding Y., Li Z., Sun Y., Li M., Shi Y. The pathogenicity of duck hepatitis A virus types 1 and 3 on ducklings. Poultry Science. 2019; 98 (12): 63336339. https://doi.org/10.3382/ps/pez455
4. Duck virus hepatitis. In: WOAH. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.3.6. https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.06_DVH.pdf
5. Huang Q., Yue H., Zhang B., Nie P., Tang C. Development of a real-time quantitative PCR for detecting duck hepatitis A virus genotype C. Journal of Clinical Microbiology. 2012; 50 (10): 3318–3323. https://doi.org/10.1128/jcm.01080-12
6. Дмитриев К. Ю. Тест-система для серологической диагностики вирусного гепатита утят типа 1 методом непрямого иммуноферментного анализа: дис. … канд. вет. наук. СПб.; 2020. 106 с.
7. Бобкова Г. Н. Инфекционные болезни птиц: учебно-методическое пособие. Брянск: ФГБОУ ВО «Брянский ГАУ»; 2015. 123 с. http://www.bgsha.com/ru/book/109816/
8. Леонов И. К. Биологические свойства вакцинных штаммов вируса гепатита утят типа I: дис. … канд. вет. наук. СПб.; 2018. 120 с.
9. Tseng C.-H., Knowles N. J., Tsai H.-J. Molecular analysis of duck hepatitis virus type 1 indicates that it should be assigned to a new genus. Virus Research. 2007; 123 (2): 190–203. https://doi.org/10.1016/j.virusres.2006.09.007
10. Fehér E., Jakab S., Bali K., Kaszab E., Nagy B., Ihász K., et al. Genomic epidemiology and evolution of duck hepatitis A virus. Viruses. 2021; 13 (8):1592. https://doi.org/10.3390/v13081592
11. Zhang R., Xia L., Chen J., Gong Y., Zhang L., Li P., et al. Molecular epidemiology and genetic diversity of duck hepatitis A virus type 3 in Shandong province of China, 2012–2014. Acta Virologica. 2017; 61 (4): 463–472. https://doi.org/10.4149/av_2017_409
12. Doan H. T. T., Le X. T. K., Do R. T., Hoang C. T. M., Nguyen K. T., Le T. H. Molecular genotyping of duck hepatitis A viruses (DHAV) in Vietnam. The Journal of Infection in Developing Countries. 2016; 10 (9): 988–995. https://doi.org/10.3855/jidc.7239
13. Hassan T. I. R., Eid A. A. M., Ghanem I. A. I., Shahin A. M., Adael S. A. A., Mohamed F. F. First report of duck hepatitis A virus 3 from duckling flocks of Egypt. Avian Diseases. 2020; 64 (3): 269–276. https://doi.org/10.1637/aviandiseases-d-19-00158
14. Фоменко В. Ю. Подбор систем культивирования возбудителя вирусного гепатита утят. Труды Федерального центра охраны здоровья животных. 2018; 16: 424–431. https://elibrary.ru/uxvpue
15. Trefilov B. B., Nikitina N. V., Yavdoshak L. I., Dmitriev K. Yu., Trubitsyn M. M. Duck hepatitis virus type 1. European Journal of Natural History. 2018; (1): 3–6. https://elibrary.ru/tbqwyh
16. Никитина Н. В., Леонов И. К., Явдошак Л. И., Трубицын М. М. Вирусный гепатит утят типа I (обзор). Птицеводство. 2023; (7–8): 74–81. https://doi.org/10.33845/0033-3239-2023-72-7-8-74-81
17. Zhang R., Yang Y., Lan J., Xie Z., Zhang X., Jiang S. Evidence of possible vertical transmission of duck hepatitis A virus type 1 in ducks. Transboundary and Emerging Diseases. 2021; 68 (2): 267–275. https://doi.org/10.1111/tbed.13708
18. Zhou S., Li S., Wang Y., Li X., Zhang T. Duck hepatitis A virus prevalence in mainland China between 2009 and 2021: a systematic review and meta-analysis. Preventive Veterinary Medicine. 2022; 208:105730. https://doi.org/10.1016/j.prevetmed.2022.105730
19. Yehia N., Erfan A. M., Omar S. E., Soliman M. A. Dual circulation of duck hepatitis A virus genotypes 1 and 3 in Egypt. Avian Diseases. 2020; 65 (1): 1–9. https://doi.org/10.1637/aviandiseases-D-20-00075
20. Rajendran R., Srinivasan J., Natarajan J., Govindan K., Kumaragurubaran K., Muthukrishnan M., et al. First report of duck hepatitis A virus genotype 2 in India. Veterinary Research Communications. 2023; 47 (3): 1231–1241. https://doi.org/10.1007/s11259-022-10063-0
21. Россельхознадзор. Инфекционные болезни животных по данным ВОЗЖ 2024.https://fsvps.gov.ru/files/infekcionnye-bolezni-zhivotnyh-po-dannym-vozzh-2024
22. Kim S. W., Yu C. D., Park J. Y., Ma X. L., Zhu T., Li Y. F., et al. The impact of genetic variation on duck hepatitis A virus (DHAV) vaccine efficacy: a comparative study of DHAV-1 and DHAV-3 against emerging variant strains. Vaccines. 2024; 12 (12):1416. https://doi.org/10.3390/vaccines12121416
23. Mao S., Ou X., Zhu D., Chen S., Ma G., Wang M., et al. Development and evaluation of indirect ELISAs for the detection of IgG, IgM and IgA1 against duck hepatitis A virus 1. Journal of Virological Methods. 2016; 237: 79–85. https://doi.org/10.1016/j.jviromet.2016.08.019
24. Liu M., Zhang T., Zhang Y., Meng F., Li X., Hou Z., et al. Development and evaluation of a VP1-ELISA for detection of antibodies to duck hepatitis type 1 virus. Journal of Virological Methods. 2010;169 (1): 66–69. https://doi.org/10.1016/j.jviromet.2010.06.018
25. Shen Y., Cheng A., Wang M., Chen S., Jia R., Zhu D., et al. Development of an indirect ELISA method based on the VP3 protein of duck hepatitis A virus type 1 (DHAV-1) for dual detection of DHAV-1 and DHAV-3 antibodies. Journal of Virological Methods. 2015; 225: 30–34. https://doi.org/10.1016/j.jviromet.2015.08.016
26. Chen L.-L., Xu Q., Zhang R.-H., Yang L., Li J.-X., Xie Z.-J., et al. Improved duplex RT-PCR assay for differential diagnosis of mixed infection of duck hepatitis A virus type 1 and type 3 in ducklings. Journal of Virological Methods. 2013; 192 (1–2): 12–17. https://doi.org/10.1016/j.jviromet.2013.04.012
27. Hu Q., Zhu D., Ma G., Cheng A., Wang M., Chen S., et al. A one-step duplex rRT-PCR assay for the simultaneous detection of duck hepatitis A virus genotypes 1 and 3. Journal of Virological Methods. 2016; 236: 207–214. https://doi.org/10.1016/j.jviromet.2016.07.011
28. Yun T., Ni Z., Hua J., Ye W., Chen L., Zhang S., et al. Development of a one-step real-time RT-PCR assay using a minor-groove-binding probe for the detection of duck Tembusu virus. Journal of Virological Methods. 2012; 181 (2): 148–154. https://doi.org/10.1016/j.jviromet.2012.01.019
29. Kim M. C., Kwon Y. K., Joh S. J., Kwon J. H., Lindberg A. M. Differential diagnosis between type-specific duck hepatitis virus type 1 (DHV-1) and recent Korean DHV-1-like isolates using a multiplex polymerase chain reaction. Avian Pathology. 2008; 37 (2): 171–177. https://doi.org/10.1080/03079450801918670
30. Chen X., Chen Y., Liu C., Li X., Liu H., Yin X., et al. Improved one-tube RTPCR method for simultaneous detection and genotyping of duck hepatitis A virus subtypes 1 and 3. PLoS ONE. 2019; 14 (8):e0219750. https://doi.org/10.1371/journal.pone.0219750
31. Chen L., Ma M., Zhang R., Xu Q., Si X., Wang Y., et al. Simultaneous detection of duck hepatitis A virus types 1 and 3, and of duck astrovirus type 1, by multiplex RT-PCR. Virologica Sinica. 2014; 29 (3): 196–198. https://doi.org/10.1007/s12250-014-3444-8
32. Lin S.-L., Cong R.-C., Zhang R.-H., Chen J.-H., Xia L.-L., Xie Z.-J., et al. Circulation and in vivo distribution of duck hepatitis A virus types 1 and 3 in infected ducklings. Archives of Virology. 2016; 161 (2): 405–416. https://doi.org/10.1007/s00705-015-2648-z
33. Wang Y., Zhu S., Hong W., Wang A., Zuo W. A multiplex PCR for detection of six viruses in ducks. Journal of Virological Methods. 2017; 248: 172–176. https://doi.org/10.1016/j.jviromet.2017.07.004
34. Li K. P., Ou S. C., Shien J. H., Chang P. C. Detection and differentiation of the vaccine strain and field isolates of duck hepatitis a virus type 1 using real-time RT-PCR and high-resolution melting assays. Taiwan Veterinary Journal. 2015; 41 (04): 1–7. https://doi.org/10.1142/S1682648515500146
35. Mao S., Wang M., Ou X., Sun D., Cheng A., Zhu D., et al. Virologic and immunologic characteristics in mature ducks with acute duck hepatitis A virus 1 infection. Frontiers in Immunology. 2017; 8:1574. https://doi.org/10.3389/fimmu.2017.01574
36. Zou Z., Ma J., Huang K., Chen H., Liu Z., Jin M. Live attenuated vaccine based on duck enteritis virus against duck hepatitis a virus types 1 and 3. Frontiers in Microbiology. 2016; 7:1613. https://doi.org/10.3389/fmicb.2016.01613
37. Roh J.-H., Kang M. Live attenuated duck hepatitis virus vaccine in breeder ducks: protective efficacy and kinetics of maternally derived antibodies. Veterinary Microbiology. 2018; 219: 107–112. https://doi.org/10.1016/j.vetmic.2018.04.021
38. Tsai H.-J. Duck hepatitis. In: Diseases of Poultry. Ed. by D. E. Swayne. 14th ed. Hoboken: Wiley-Blackwell; 2020; 450–459.
39. Князев В. П. Болезни водоплавающих птиц. Владимир; Покров; 2011. 327 с.
40. Вакцина против вирусного гепатита утят из штамма «ВГНКИ-К» эмбриональная. https://shop.arriah.ru/catalog/vaktsiny/vaktsiny-protiv-bolezney-ptits/gepatit-utyat-zhid-shtvgnkik/?ysclid=mf11a894fp704916994
41. AVAC DVH Live (Duck viral hepatitis Vaccine). https://www.avac.com.vn/en/products-for-poultry/avacdvhlive
42. Zhang Y., Wu S., Liu W., Hu Z. Current status and future direction of duck hepatitis A virus vaccines. Avian Pathology. 2023; 52 (2): 89–99. https://doi.org/10.1080/03079457.2022.2162367
43. Kim M.-C., Kim M.-J., Kwon Y.-K., Lindberg A. M., Joh S.-J., Kwon H.-M., et al. Development of duck hepatitis A virus type 3 vaccine and its use to protect ducklings against infections. Vaccine. 2009; 27 (48): 6688–6694. https://doi.org/10.1016/j.vaccine.2009.08.092
44. Wu F., Lu F., Fan X., Pan Q., Zhao S., Sun H., et al. Development of a live attenuated duck hepatitis A virus type 3 vaccine (strain SD70). Vaccine. 2020; 38 (30): 4695–4703. https://doi.org/10.1016/j.vaccine.2020.05.030
45. Kang M., Roh J.-H., Jang H.-K. Protective efficacy of a bivalent live attenuated vaccine against duck hepatitis A virus types 1 and 3 in ducklings. Veterinary Microbiology. 2018; 214: 108–112. https://doi.org/10.1016/j.vetmic.2017.12.018
46. Gough R. E., Spackman D. Studies with inactivated duck virus hepatitis vaccines in breeder ducks. Avian Pathology. 1981. 10 (4): 471–479. https://doi.org/10.1080/03079458108418497
47. Liu X., Kong X. Isolation, identification and attenuation of a pathogenic duck hepatitis virus type 1 in China, and complete genomic sequence comparison between the embryo-passaged, attenuated derivatives and their parent. Polish Journal of Veterinary Sciences. 2019; 22 (1): 163–171. https://doi.org/10.24425/pjvs.2018.125614
48. Wang W., Said A., Wang B., Qu G., Xu Q., Liu B., Shen Z. Establishment and evaluation of the goose embryo epithelial (GEE) cell line as a new model for propagation of avian viruses. PloS ONE. 2018; 13 (3):e0193876. https://doi.org/10.1371/journal.pone.0193876
49. Wang M., Li Z., Liu H., Wang X., Zhang D. Effect of fetal calf serum on propagation of duck hepatitis A virus genotype 3 in duck embryo fibroblast cells. BMC Veterinary Research. 2019; 15 (1):153. https://doi.org/10.1186/s12917-019-1904-y
50. Wang M., Chai L., Liang S., Lv J., Yang L., Qu S., et al. Fetal calf serum exerts an inhibitory effect on replication of duck hepatitis A virus genotype 1 in duck embryo fibroblast cells. Viruses. 2020; 12 (1):80. https://doi.org/10.3390/v12010080
51. Никитина Н. В., Явдошак Л. И., Леонов И. К., Трубицын М. М. Профилактика вирусного гепатита утят типа I и меры борьбы: методические положения. СПб.: ВНИВИП; 2024. 15 с. https://elibrary.ru/fliwda
52. Трефилов Б. Б., Никитина Н. В., Леонов И. К. Кинетика инактивации вируса гепатита утят типа I (Avihepatovirus, Picornaviridae). Вопросы вирусологии. 2018; 63 (3): 135–138. https://doi.org/10.18821/0507-4088-2018-63-3-135-138
53. Трефилов Б. Б., Никитина Н. В., Явдошак Л. И., Трубицын И. И. Инактивированная эмульгированная вакцина против вирусного гепатита утят типа I. Ветеринария. 2018; (2): 20–23. https://elibrary.ru/yqbtfm
54. Трубицын М. М., Никитина Н. В. Опыт применения экспериментальной инактивированной вакцины против вирусного гепатита утят типа I. Актуальные проблемы лечения и профилактики болезней молодняка: материалы Международной научно-практической конференции (Витебск, 2–4 ноября 2023 г.). Витебск: УО ВГАВМ; 2023; 386-389. https://elibrary.ru/urnpcf
55. Никитина Н. В., Трубицын М. М. Опыт применения инактивированной вакцины против вирусного гепатита утят типа I. Птицеводство. 2021; (12): 69–72. https://doi.org/10.33845/0033-3239-2021-70-12-69-72
56. Yin F., Li J., Zhang S., Yu M., Zhang W., Fan G., et al. Development and evaluation of an inactivated bivalent vaccine against duck viral hepatitis. Chinese Journal of Biotechnology. 2015; 31 (11): 1579–1588. https://doi.org/10.13345/j.cjb.140636 (in Chinese)
57. Zou Z., Hu Y., Liu Z., Zhong W., Cao H., Chen H., Jin M. Efficient strategy for constructing duck enteritis virus-based live attenuated vaccine against homologous and heterologous H5N1 avian influenza virus and duck enteritis virus infection. Veterinary Research. 2015; 46:42. https://doi.org/10.1186/s13567-015-0174-3
58. Niu Y., Liu B., Sun C., Zhao L., Chen H. Construction of the recombinant duck enteritis virus delivering capsid protein VP0 of the duck hepatitis A virus. Veterinary Microbiology. 2020; 249:108837. https://doi.org/10.1016/j.vetmic.2020.108837
59. Yang F., Liu P., Li X., Liu R., Gao L., Cui H., et al. Recombinant duck enteritis virus-vectored bivalent vaccine effectively protects against duck hepatitis A virus infection in ducks. Frontiers in Microbiology. 2021; 12:813010. https://doi.org/10.3389/fmicb.2021.813010
60. Wang A. P., Liu L., Gu L. L., Guo C. M., Wu S., Feng Q., et al. Protection against duck hepatitis A virus type 1 conferred by a recombinant avian adeno-associated virus. Poultry Science. 2019; 98 (1): 112–118. https://doi.org/10.3382/ps/pey325
61. Wang A. P., Liu L., Gu L. L., Wu S., Guo C. M., Feng Q., et al. Expression of duck hepatitis A virus type 1 VP3 protein mediated by avian adeno-associated virus and its immunogenicity in ducklings. Acta Virologica. 2019; 63 (1): 53–59. https://doi.org/10.4149/av_2019_104
62. Zheng W. Q., Song M. X., Li J. L., Huang B., Li Y. F., Yu K. X., et al. Generation and evaluation of recombinant Newcastle disease viruses (NDV) expressing VP1 gene of duck hepatitis A viruses (DHAV) type 1 and 3. Chinese Journal of Veterinary Science. 2017; 37 (9): 1641–1647. https://www.cabidigitallibrary.org/doi/full/10.5555/20173348071 (in Chinese)
63. Li X., Zhao R., Lin W., Li C., Zhang T., Meng F., et al. Evidence of VP1 of duck hepatitis A type 1 virus as a target of neutralizing antibodies and involving receptor-binding activity. Virus Research. 2017; 227: 240–244. https://doi.org/10.1016/j.virusres.2016.10.018
64. Wang C., Li X. K., Wu T. C., Wang Y., Zhang C. J., Cheng X. C., Chen P. Y. Recombinant VP1 protein of duck hepatitis virus 1 expressed in Pichia pastoris and its immunogenicity in ducks. Acta Virologica. 2014; 58 (4): 333–339. https://doi.org/10.4149/av_2014_04_333
65. Truong T.-N., Cheng L.-T. Development of a subunit vaccine against duck hepatitis A virus serotype 3. Vaccines. 2022; 10 (4):523. https://doi.org/10.3390/vaccines10040523
66. Zell R., Delwart E., Gorbalenya A. E., Hovi T., King A. M. Q., Knowles N. J., et al. ICTV virus taxonomy profile: Picornaviridae. Journal of General Virology. 2017; 98 (10): 2421–2422. https://doi.org/10.1099/jgv.0.000911
67. Ku Z., Ye X., Huang X., Cai Y., Liu Q., Li Y., et al. Neutralizing antibodies induced by recombinant virus-like particles of enterovirus 71 genotype C4 inhibit infection at pre- and post-attachment steps. PLoS ONE. 2013; 8 (2):e57601. https://doi.org/10.1371/journal.pone.0057601
68. Somasundaram B., Chang C., Fan Y. Y., Lim P.-Y., Cardosa J., Lua L. Characterizing Enterovirus 71 and Coxsackievirus A16 virus-like particles production in insect cells. Methods. 2016; 95: 38–45. https://doi.org/10.1016/j.ymeth.2015.09.023
69. Marsian J., Fox H., Bahar M. W., Kotecha A., Fry E. E., Stuart D. I., et al. Plantmade polio type 3 stabilized VLPs-a candidate synthetic polio vaccine. Nature Communications. 2017; 8 (1):245. https://doi.org/10.1038/s41467-017-00090-w
70. Wang A., Gu L., Wu S., Zhu S. Duck hepatitis A virus structural proteins expressed in insect cells self-assemble into virus-like particles with strong immunogenicity in ducklings. Veterinary Microbiology. 2018; 215: 23–28. https://doi.org/10.1016/j.vetmic.2017.12.020
71. Гален. Государственный реестр лекарственных средств для ветеринарного применения. https://galen.vetrf.ru/#/registry/pharm/registry?page=1
Куникова Екатерина Дмитриевна, ведущий технолог лаборатории профилактики болезней птиц
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Волков Михаил Сергеевич, д-р вет. наук, доцент, заведующий лабораторией эпизоотологии и мониторинга
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Мороз Наталья Владимировна, канд. вет. наук, заведующий лабораторией профилактики болезней птиц
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901

|
1. Таблица 2. Сравнительный анализ профилей эффективности вакцинных платформ | |
| Тема | ||
| Тип | Исследовательские инструменты | |
Скачать
(456KB)
|
Метаданные ▾ | |
Куникова Е.Д., Волков М.С., Мороз Н.В. Специфическая профилактика вирусного гепатита утят (обзор). Ветеринария сегодня. 2026;15(2):110-122. https://doi.org/10.29326/2304-196X-2026-15-2-110-122
Kunikova E.D., Volkov M.S., Moroz N.V. Specific prevention of duck virus hepatitis (review). Veterinary Science Today. 2026;15(2):110-122. (In Russ.) https://doi.org/10.29326/2304-196X-2026-15-2-110-122
600901, Владимирская область, город Владимир, микрорайон Юрьевец, улица Гвардейская, дом 6
ФГБУ «ВНИИЗЖ»
тел.: 8 (4922) 26-15-12, доб. 22-27
E-mail: nikeshina@arriah.ru






























