




Содержание
Перейти к:
https://doi.org/10.29326/2304-196X-2026-15-2-177-183
Перейти к:
Введение. Ньюкаслская болезнь регистрируется во многих странах мира, иногда принимая характер эпизоотии. Активно циркулирующий в последние годы вирус ньюкаслской болезни генотипа VII отличается высокой вирулентностью и вызывает беспокойство в силу способности активно мутировать. В условиях интенсивного птицеводства важно уделять особое внимание вопросам специфической профилактики данной болезни и применять эффективные вакцины для защиты птицепоголовья.
Цель исследования. Определить иммуногенную активность трех инактивированных вакцин против ньюкаслской болезни птиц при контрольном заражении цыплят вирусом ньюкаслской болезни генотипа VII.
Материалы и методы. Исследованы три вакцины против ньюкаслской болезни: моновалентная (антиген «Ла-Сота»), «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти» (антигены «ВНИИЗЖ G7» и «Ла-Сота»). Антигены разбавляли физиологическим раствором в соотношениях 1:25, 1:50, 1:100 или использовали неразведенными, затем эмульгировали с адъювантом Coralvac RZ 528. Каждый образец соответствующей вакцины вводили 10 цыплятам яичного кросса 4-недельного возраста внутримышечно в область груди в объеме 0,5 см3. Контрольную группу не вакцинировали. Через 28 сут определяли титр поствакцинальных антител в реакции торможения гемагглютинации и проводили контрольное заражение вирулентным штаммом вируса ньюкаслской болезни генотипа VII.
Результаты. Все исследуемые препараты вызывали формирование напряженного поствакцинального иммунитета к вирусу ньюкаслской болезни спустя 28 сут после введения птицам, а также соответствовали требованиям Всемирной организации здравоохранения животных. Тем не менее вакцины «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти» показали более высокий уровень протективного действия при контрольном заражении изолятом вируса ньюкаслской болезни генотипа VII в сравнении с моновалентной вакциной из антигена «Ла-Сота».
Заключение. Поливалентные вакцины «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти», изготовленные на основе смеси антигенов вируса ньюкаслской болезни генотипов II и VII, обеспечивают высокую степень защиты цыплят и могут использоваться для профилактики ньюкаслской болезни, вызванной вирусами генотипа VII.
Вершинина М.А., Мороз Н.В., Фролов С.В. Оценка иммуногенного действия вакцин против ньюкаслской болезни. Ветеринария сегодня. 2026;15(2):177-183. https://doi.org/10.29326/2304-196X-2026-15-2-177-183
Vershinina M.A., Moroz N.V., Frolov S.V. Assessment of immunogenic activity of Newcastle disease vaccines. Veterinary Science Today. 2026;15(2):177-183. https://doi.org/10.29326/2304-196X-2026-15-2-177-183
Ньюкаслская болезнь (НБ) представляет собой высококонтагиозное вирусное заболевание, которое поражает различные виды птиц в зависимости от степени их восприимчивости [1] и наносит огромный экономический ущерб птицеводческой отрасли [2].
Первые случаи НБ птиц были зафиксированы в 1926 г. в Индонезии и в 1927 г. в Ньюкасл-апон-Тайне (Англия) [3]. С тех пор о крупных вспышках НБ сообщалось во многих странах, включая Корею, Филиппины, Индию, Шри-Ланку и Японию [4].
Клинические признаки болезни нередко отличаются в зависимости от вида птицы и конкретного штамма возбудителя. Известно, что куры крайне восприимчивы, в то время как утки и гуси менее подвержены действию вируса [5]. Возбудитель способен поражать более 236 видов птиц, относящихся как минимум к 20 отрядам, при этом могут быть инфицированы как домашние, так и дикие птицы [6][7]. У домашней птицы характерными симптомами являются потеря аппетита, угнетение, аномальная жажда, слабость, отеки и снижение яйценоскости; респираторные проявления могут включать чихание, затрудненное дыхание, выделения из носа и кашель, в то время как явным кишечным симптомом является диарея с пометом зеленоватого цвета [8]. К поражениям нервной системы относятся кривошея, парезы, параличи, атаксия, а также тремор головы и конечностей. При остром течении НБ смерть наступает внезапно, птицы могут погибать без каких-либо клинических признаков [8].
Возбудителем болезни является представитель рода Orthoavulavirus подсемейства Avulavirinae семейства Paramyxoviridae [2]. Это оболочечный вирус, генетический материал которого представлен несегментированной негативно-смысловой одноцепочечной молекулой РНК длиной приблизительно 15 000 нуклеотидов [9][10]. Ключевым фактором патогенности возбудителя ньюкаслской болезни является образование активных слитых белков F1 и F2 при расщеплении предшественника F-белка (F0), а также наличие некоторых основных остатков в месте расщепления слитого белка (FPCS) [11].
С момента своего появления вирус НБ птиц претерпел заметную эволюцию, что проявилось в значительном генетическом, вирулентном и антигенном разнообразии, а также в расширении ареала распространения [12]. Филогенетический анализ последовательности гена белка F позволяет разделить штаммы вируса НБ на два класса [13]. Класс I включает преимущественно авирулентные штаммы вируса, природным резервуаром которых являются водоплавающие дикие птицы, в то время как к классу II относятся варианты вируса с более высокой генетической изменчивостью и вирулентностью по меньшей мере 20 генотипов, которые инфицируют различные виды домашних и диких птиц [14]. Кроме того, штаммы вируса класса II считаются ответственными за большинство вспышек НБ, о которых известно на сегодняшний день [15]. Генотипы II, III и IV были ответственны за первые эпизоотии НБ, которые произошли с 1920 по 1960 г., а субгенотип VIb (субгенотип VI.1.1, согласно современной классификации) привел к третьей панзоотии среди голубей в 1980-х гг. [16]. Генотип VII, в свою очередь, вызвал четвертую панзоотию НБ, охватившую Азию, Африку, Европу и Южную Америку [2], и пятую, последнюю на данный момент, панзоотию [16]. Варианты вируса НБ генотипа VII считаются наиболее быстро эволюционирующими в пределах своего класса [17], а их высокая вирулентность и способность вызывать болезнь даже у водоплавающих птиц оправданно вызывает беспокойство в мировом сообществе [18].
Поскольку эффективность каких-либо лекарственных препаратов в лечении ньюкаслской болезни птиц не доказана, единственным способом защиты стада является профилактика, которая достигается путем соблюдения мер биозащиты и вакцинации [19].
На коммерческих птицефабриках основными биологическими рисками являются неправильная утилизация мертвой птицы и неполное обеззараживание помета, совместное содержание разновозрастного и многовидового поголовья птицы, а также ненадлежащий контроль за перемещениями и санитарией на птицефабрике [20]. В связи с этим основным направлением биозащиты является минимизация контакта домашней птицы, кормов, воды и рабочего инвентаря с любыми факторами, расположенными за пределами птицефабрики, особенно с дикими птицами.
Вакцинация лежит в основе стратегии борьбы с ньюкаслской болезнью и защиты популяций домашних птиц в эндемичных регионах [21]. Наиболее часто используемые вакцины против НБ разработаны на основе живых или инактивированных штаммов вируса, выделенных в прошлом веке, включая штаммы LaSota, B1, Ulster/67, Mukteswar и VG/GA [22]. Хотя современные вакцины при должном применении могут предотвратить вспышку болезни и гибель поголовья, они не всегда препятствуют распространению вируса [23]. Потенциальное антигенное несоответствие между вакцинными и полевыми штаммами способно отразиться на эффективности иммунопрофилактики. В этих условиях полевые изоляты вируса все еще могут распространяться среди птиц и вызывать атипичное течение болезни [24]. Имеющиеся данные свидетельствуют о том, что вакцины на основе циркулирующих штаммов демонстрируют более высокий уровень защиты [25] и способствуют снижению выделения вируса по сравнению с вакцинами, изготовленными из штаммов, сильно отличающихся от полевых [24]. Также сообщалось, что значительная генетическая разница между циркулирующими в настоящее время полевыми изолятами генотипа VII и вакцинными штаммами генотипа I или II играет критическую роль в частой подверженности птицефабрик вспышкам НБ, обусловленным вирулентным вирусом, несмотря на применяемые стратегии вакцинации [26].
Вышеуказанные данные подчеркивают необходимость усовершенствования мер специфической профилактики ньюкаслской болезни, в частности актуализации производственных вакцинных штаммов вируса НБ. Целью исследования являлось изучение иммуногенной активности моновалентной вакцины на основе традиционно применяемого антигена вируса НБ генотипа II, а также вакцин «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти», изготовленных с использованием антигенов вируса НБ II и VII генотипов.
Объекты исследования:
1) вакцина против ньюкаслской болезни инактивированная эмульсионная (далее – «НБ-Ла-Сота»); активный компонент – антиген вируса НБ штамма «Ла-Сота» в составе экстраэмбриональной жидкости (ЭЭЖ) инфицированных эмбрионов кур;
2) вакцина против ньюкаслской болезни инактивированная эмульсионная «ВНИИЗЖ-АвиНью Мульти»; активный компонент – антиген вируса НБ, состоящий из смеси штаммов «Ла-Сота» и «ВНИИЗЖ G7» в составе ЭЭЖ инфицированных эмбрионов кур;
3) вакцина против ньюкаслской болезни («G7» и «Ла-Сота») и гриппа птиц H9N2 (Y280 и G1) ассоциированная инактивированная эмульсионная «ВНИИЗЖ-АвиНьюФлу Мульти»; активный компонент – антиген вируса НБ, состоящий из смеси штаммов «Ла-Сота» и «ВНИИЗЖ G7» в составе ЭЭЖ инфицированных эмбрионов кур.
Производственные штаммы вирусов. В эксперименте использовали:
– штамм «ВНИИЗЖ G7» вируса НБ генотипа VII субгенотипа VII.1.1. (далее – «ВНИИЗЖ G7»);
– штамм «Ла-Сота» вируса НБ генотипа II (далее – «Ла-Сота»);
– штамм «A/chicken/Amursky/03/12/H9N2» вируса низкопатогенного гриппа птиц подтипа H9N2;
– штамм «Челябинск-20» вируса низкопатогенного гриппа птиц подтипа H9N2.
Вирусы инактивировали аминоэтилэтиленимином (0,25%). Специфическая активность полученных антигенов вируса НБ составила: 7 log2 (1:128) ГАЕ – для «ВНИИЗЖ G7» и 10 log2 (1:1024) ГАЕ – для «Ла-Сота». Специфическая инфекционная активность вирусов до инактивации: 9,0 ЭИД50/см³ – для «ВНИИЗЖ G7»; 10,5 ЭИД50/см³ – для «Ла-Сота».
Получение разведений вакцин с заданной концентрацией антигена. В качестве исходного антигена вируса НБ для изготовления вакцины «НБ-Ла-Сота» использовали цельный (неразведенный) антиген штамма «Ла-Сота», а для изготовления вакцин «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти» – смесь антигенов штаммов «Ла-Сота» и «ВНИИЗЖ G7» в равном соотношении. Концентрации антигенов (D) в прививном объеме вакцин регулировали путем разведения исходных препаратов соответствующих антигенов физиологическим раствором в соотношениях 1:25, 1:50 и 1:100, а также применяли неразведенные антигены.
Далее полученные препараты антигенов эмульгировали в соотношении 30:70 в масляном адъюванте Coralvac RZ 528 (Турция).
При изготовлении разведений вакцины «ВНИИЗЖ-АвиНьюФлу Мульти» использовали также антигены вируса низкопатогенного гриппа птиц подтипа H9N2 (штаммы «A/chicken/Amursky/03/12/H9N2» и «Челябинск-20»).
Указанные антигены вирусов НБ и гриппа птиц использовали в соотношении 1:1, при этом количество антигенов разных штаммов было в равных пропорциях.
Смешивание с разбавителем производили в лабораторном гомогенизаторе тканей при 6000 оборотов в течение 10 мин. Стабильность полученной эмульсии оценивали центрифугированием при 1000 g в течение 10 мин. Эмульсию считали стабильной, если отслоение легкой (масляной) фракции не превышало 5% по объему, а отслоения тяжелой (водной) фракции не происходило.
Птица. Исследование проводили на 4-недельных цыплятах яичного кросса Ломан Браун из хозяйства, благополучного по острым инфекционным болезням, серонегативных к вирусу НБ. Отсутствие специфических антител к вирусу НБ подтверждали путем исследования сывороток крови, полученных до заражения птиц, в реакции торможения гемагглютинации (РТГА).
Иммунизация птиц. Каждый образец вакцины с заданной концентрацией антигена был испытан на отдельной группе птиц численностью 10 гол. Препарат вводили внутримышечно в область груди в объеме 0,5 см³. Дополнительно была образована группа контроля вируса (10 гол.), в которой иммунизацию не проводили (интактные птицы). Опытные группы формировались в соответствии с «Правилами регулирования обращения ветеринарных лекарственных средств на таможенной территории Евразийского экономического союза» (утверждены решением Совета Евразийской экономической комиссии от 21.01.2022 № 1)1. Птиц содержали группами в изолированных настольных боксах с автономной вентиляцией, подачей воды и корма.
Реакция гемагглютинации (РГА). Пробы антигенсодержащих материалов исследовали в РГА в соответствии с методикой, изложенной в Руководстве по диагностическим тестам и вакцинам для наземных животных Всемирной организации здравоохранения животных (Руководство ВОЗЖ)2. Определяли титр гемагглютинирующих единиц (ГАЕ).
Реакция торможения гемагглютинации (РТГА). C помощью РТГА в сыворотках крови, полученных от птиц через 28 сут после вакцинации, определяли титр антител к вирусу НБ, используя диагностические наборы производства ФГБУ «ВНИИЗЖ», в соответствии с «Методическими рекомендациями по идентификации вирусов гриппа птиц и ньюкаслской болезни в реакции торможения гемагглютинации»3.
Заражение птиц. В группах иммунизированных и интактных птиц проводили контрольное заражение через 28 сут после вакцинации, для чего использовали вирулентный изолят вируса НБ NDV/chicken/rus/Saratov/2403-3/22. Вирусный материал вводили в дозе 6,0 lgЭИД50/0,5 см³ внутримышечно в область бедра.
В течение последующих 10 сут оценивали состояние зараженной и контактной птицы согласно ГОСТ Р 58090-2018 «Клиническое обследование непродуктивных животных. Общие требования»4. Проводили ежедневное наблюдение за клиническим состоянием опытных птиц с фиксацией фаз течения болезни, различных проявлений и признаков болезни, а также гибели.
По окончании срока наблюдений в группах определяли суммарное число клинически больных и павших птиц, зафиксированное за период опыта (∑c), на основании которого вычисляли протективные индексы (Pr) по формуле:
Pr = 1 – (∑c) / n,
где n – количество птиц в группе до заражения.
Все эксперименты на животных проводились в строгом соответствии с межгосударственным стандартом по содержанию и уходу за лабораторными животными ГОСТ 33215-2014, принятым Межгосударственным советом по стандартизации, метрологии и сертификации, а также согласно требованиям Директивы 2010/63/EU Европейского парламента и Совета Европейского союза от 22.09.2010 по охране животных, используемых в научных целях. Исследования одобрены комиссией по биоэтике ФГБУ «ВНИИЗЖ» (заключение от 26.09.2025).
Показатели напряженности поствакцинального иммунитета. Оценками напряженности поствакцинального гуморального иммунитета к вирусу НБ служили установленные в РТГА средние логарифмические титры антител к вирусу НБ (Т). Для постановки реакции параллельно использовали антигены «ВНИИЗЖ G7» и «Ла-Сота». Результаты клинических наблюдений в группах после контрольного заражения выражали в виде клинических показателей и индексов защиты.
Обработка экспериментальных данных. Использовали общепринятые способы обработки выборок варьирующих переменных (определяли средние значения, стандартные отклонения и стандартные ошибки средних). Применяли регрессионный анализ [27]. Для приближения зависимости между величинами к линейному виду значения D (величина разведения антигена) логарифмировали (lgD), при этом оценки Pr преобразовывали в линейные эквиваленты. Линейные эквиваленты вычисляли с помощью логит-преобразования по Берксону [28]. Для эмпирических значений Р = 1 использовали условные оценки:
‘Р1 = 1 – (1 / 5n) [29].
Расчет величины 50%-й протективной дозы (PD50) выполняли по формуле:
lgPD50 = b / (–k),
где b – свободный член регрессионного уравнения (ордината линии регрессии);
k – регрессионный коэффициент.
Статистическую неопределенность показателя lgPD50 [30] указывали в виде стандартной ошибки измерения (±S).
За величину PD50, согласно Руководству ВОЗЖ, принимали протективный эффект вакцины в разведении, при котором наблюдается 50%-я защита животных. Вакцину считали соответствующей стандарту, если значение PD50 составляло не менее 50 на дозу и если нижний доверительный предел был не менее 35 PD50 на дозу5.
Вычислительные операции и графические построения выполняли в приложении Excel.
В ходе проведенного эксперимента было установлено, что исследуемые вакцины против ньюкаслской болезни птиц обладают различными иммуногенными свойствами в отношении вирулентного изолята вируса НБ генотипа VII. В таблице 1 представлены первичные данные по изучению протективной активности вакцин против НБ. Даны разведения антигена в прививном объеме вакцины, испытанные на группах из n-числа птиц, протективные индексы (Pr) и соответствующие им линейные эквиваленты (Y).
Таблица 1
Оценки протективного эффекта, установленные для трех инактивированных вакцин, содержащих заданные дозы антигенов «ВНИИЗЖ G7» и «Ла-Сота» вируса НБ, после контрольного заражения изолятом NDV/chicken/rus/Saratov/2403-3/22
Table 1
Protective efficacy estimates for three inactivated vaccines containing specified doses of NDV ARRIAH G7 and LaSota antigens after challenge with the NDV/chicken/rus/Saratov/2403-3/22 isolate
Вакцина | D* (lgD) | n | ∑c** | Pr = 1 – (∑c) / n | Y = log (Pr / (1 – Pr)) |
«ВНИИЗЖ-АвиНью Мульти» | 1 (0) | 10 | 0 | 0,98# | 1,690 |
25 (1,4) | 10 | 1 | 0,90 | 0,954 | |
50 (1,7) | 10 | 3 | 0,70 | 0,368 | |
100 (2,0) | 10 | 5 | 0,50 | 0,000 | |
«ВНИИЗЖ-АвиНьюФлу Мульти» | 1 (0) | 10 | 0 | 0,98 | 1,690 |
25 (1,4) | 10 | 1 | 0,90 | 0,954 | |
50 (1,7) | 10 | 3 | 0,70 | 0,368 | |
100 (2,0) | 10 | 6 | 0,40 | –0,176 | |
«НБ-Ла-Сота» | 1 (0) | 10 | 1 | 0,90 | 0,954 |
25 (1,4) | 10 | 3 | 0,70 | 0,368 | |
50 (1,7) | 10 | 4 | 0,60 | 0,176 | |
100 (2,0) | 10 | 6 | 0,40 | –0,176 | |
Контроль | – | 10 | 10 | – | – |
* указана величина разведения антигена в прививном объеме вакцины (D = 1 соответствует неразведенному антигену);
** число клинически больных и павших птиц, зафиксированное в группе в период опыта;
# для эмпирических оценок Pr = 1 – (∑c) / n = 1 принята условная величина “Pr1 = 1 – (1 / 5n).
* the antigen dilution value in the vaccine inoculation volume is indicated (D = 1 corresponds to undiluted antigen);
** the number of clinically diseased and dead birds recorded in the group during the experimental period;
# for empirical estimates of Pr = 1 – (∑c) / n = 1, a conditional value of Pr1 = 1 – (1 / 5n) was taken.
Соответственно тестируемым препаратам исследовали зависимость значений эквивалентов (Y) от величин испытанных прививных доз антигенов (D). Построили регрессионные модели, объективно отражающие связь исследуемых параметров с учетом общей статистики всех первичных данных, и определяли показатели PD50, характеризующие протективные потенциалы тестируемых вакцин.
Результаты регрессионного анализа в графическом виде представлены на рисунке.
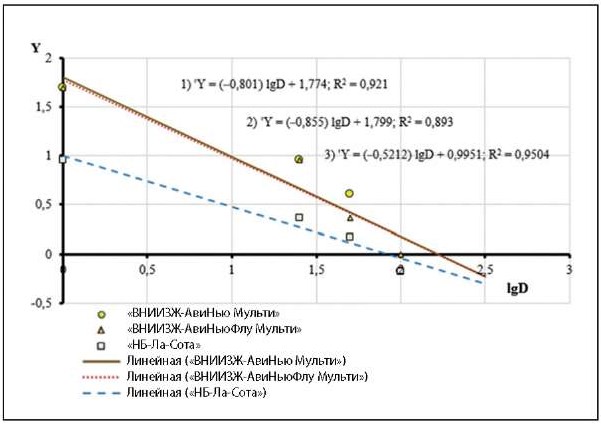
Рис. Зависимость между испытанными разведениями антигенов (lgD), входящими в состав вакцин, и эквивалентами индекса защиты (Y) иммунизированных птиц после контрольного заражения изолятом NDV/chicken/rus/Saratov/2403-3/22
Примечание: Показано положение экспериментальных оценок Y по оси lgD. Ось Y пересечена в точке Y = 0, соответствующея эффекту 50%-й защиты. Приведены регрессионные уравнения (модели), где ‘Y – ожидаемая величина эквивалента индекса защиты для заданного значения lgD: 1) «ВНИИЗЖ-АвиНью Мульти»; 2) «ВНИИЗЖ-АвиНьюФлу Мульти»; 3) «НБ-Ла-Сота». Даны коэффициенты адекватности моделей (R²), характеризующие степень соответствия их экспериментальным точкам.
Fig. Relationship between the tested antigen dilutions (lgD) included in the vaccines and the protection index equivalents (Y) of immunized birds after challenge with the NDV/chicken/rus/Saratov/2403-3/22 isolate
Note: The position of experimental Y estimates along the lgD axis is indicated. The Y-axis is intersected at the point Y = 0, which correspons to the 50% protective effect. Regression equations (models) are provided, where ‘Y is the expected protection index equivalent for a given lgD value: 1) ARRIAH-AviNew Multi; 2) ARRIAH-AviNewFlu Multi; 3) ND-LaSota. The model adequacy coefficients (R²) characterizing the degree of correspondence between the models and the experimental data points are provided.
Как видно, уравнения (регрессионные модели) имели высокий уровень адекватности (R² > 0,8), что позволяло использовать их для дальнейшего вычисления концентрации 50%-х протективных доз (PD50), которые при данных условиях испытаний содержались в прививном объеме тестируемых препаратов, равном 0,5 см³. Полученные результаты представлены в таблице 2.
Таблица 2
Показатели lgPD50 ± S, установленные после контрольного заражения птиц изолятом NDV/chicken/rus/Saratov/2403-3/22 соответственно испытанным вакцинам
Table 2
The lgPD50 ± S values determined following challenge of birds with the NDV/chicken/rus/Saratov/2403-3/22 isolate for each of the tested vaccines
Вакцина | Показатель 50%-й протективной дозы, lgPD50 ± S | PD50 в прививном объеме вакцины, PD50/0,5 см³ |
«ВНИИЗЖ-АвиНью Мульти» | 1,7738/0,8006 = 2,215 ± 0,25 | 164 (92)* |
«ВНИИЗЖ-АвиНьюФлу Мульти» | 1,7991/ 0,8550 = 2,104 ± 0,32 | 127 (61) |
«НБ-Ла-Сота» | 0,9951/0,5212 = 1,909 ± 0,13 | 81 (60) |
* в скобках указан нижний интервал значения PD50/0,5 см³ для каждой из испытанных вакцин (the lower interval of PD50/0.5 cm³ for each of the tested vaccines is shown in parentheses).
В эксперименте по оценке протективного эффекта вакцин против заражения высоковирулентным изолятом NDV/chicken/rus/Saratov/2403-3/22 установлено, что наибольшим протективным потенциалом обладал препарат «ВНИИЗЖ-АвиНью Мульти», который в прививном объеме содержал 164 PD50/0,5 см³. Следующим по данному показателю был препарат «ВНИИЗЖ-АвиНьюФлу Мульти», содержавший 127 PD50/0,5 см³. Вакцина «НБ-Ла-Сота» имела наименьшую оценку, которая составила 81 PD50/0,5 см³.
Далее проводили серологические исследования. В РТГА определяли титры антител к вирусу НБ (T), установленные в пробах сыворотки крови птиц через 28 сут после иммунизации. Анализировали связь значений титров и заданных прививных доз антигенов и сравнивали значения титров, полученных в гомо- (hom) и гетерологичных (het) реакциях. Соответствующие результаты приведены в таблице 3.
Таблица 3
Результаты исследования в РТГА сывороток крови цыплят через 28 сут после иммунизации тремя инактивированными вакцинами, содержащими в заданных дозах антигены «ВНИИЗЖ G7» и «Ла-Сота» вируса НБ
Table 3
Hemagglutination inhibition test results of chicken sera in 28 days post-immunization with three inactivated vaccines containing specified doses of NDV ARRIAH G7 and LaSota antigens
Вакцина | Разведение антигена в вакцине | Титры антител к вирусу НБ соответственно использованному антигену, log2 T | |
D (lgD) | «ВНИИЗЖ G7» (hom) | «Ла-Сота» (het) | |
«ВНИИЗЖ-АвиНью Мульти» | 1 (0) | 10,0 ± 0,12 | 9,30 ± 0,11 |
25 (1,4) | 9,20 ± 0,12 | 6,80 ± 0,12 | |
50 (1,7) | 7,90 ± 0,12 | 2,80 ± 0,14 | |
100 (2,0) | 5,30 ± 0,12 | 2,50 ± 0,10 | |
«ВНИИЗЖ-АвиНьюФлу Мульти» | 1 (0) | 8,10 ± 0,10 | 6,40 ± 0,11 |
25 (1,4) | 5,40 ± 0,10 | 3,90 ± 0,10 | |
50 (1,7) | 5,40 ± 0,11 | 3,70 ± 0,12 | |
100 (2,0) | 4,40 ± 0,10 | 2,60 ± 0,10 | |
D (lgD) | «ВНИИЗЖ G7» (het) | «Ла-Сота» (hom) | |
«НБ-Ла-Сота» | 1 (0) | 7,20 ± 0,10 | 9,30 ± 0,10 |
25 (1,4) | 6,20 ± 0,12 | 8,10 ± 0,09 | |
50 (1,7) | 5,50 ± 0,08 | 7,00 ± 0,12 | |
100 (2,0) | 5,20 ± 0,09 | 6,80 ± 0,14 | |
Исходя из полученных данных можно заключить, что напряженность поствакцинального гуморального иммунитета зависела от концентрации антигена в прививном объеме вакцины. Значения титров антител в группах цыплят, привитых цельным антигеном в составе вакцины, были в пределах от 6,4 до 10,0 log2. Также было отмечено, что титры антител в РТГА несколько различались при постановке реакции с различными антигенами. Наибольшие титры антител к антигену «Ла-Сота» выявляли в группах цыплят, привитых вакцинами «ВНИИЗЖ-АвиНью Мульти» и «НБ-Ла-Сота»: (9,30 ± 0,11) и (9,30 ± 0,10) log2 соответственно. Наиболее высокие титры антител к антигену «ВНИИЗЖ G7» выявляли после вакцинации «ВНИИЗЖ-АвиНью Мульти».
Кроме того, установили, что значение титров пост-вакцинальных антител в гомологичных реакциях (log2 Thom) превышали аналогичные показатели в гетерологичных реакциях (log2 Thet), что указывало на меньшую эффективность образования иммунных комплексов в гетерологичной системе.
Таким образом, на основании результатов контрольного заражения и результатов оценки гуморального иммунитета установили, что все вакцины соответствовали требованиям ВОЗЖ, поскольку полученные значения 50%-й протективной дозы превышали 50 PD50/0,5 см³, а нижний интервал был выше 35 PD50/0,5 см³. Тем не менее среди испытуемых образцов более иммуногенным препаратом для контроля ньюкаслской болезни у кур, вызванной вирусами генотипа VII, оказалась вакцина «ВНИИЗЖ-АвиНью Мульти». Полученные в ходе исследования данные подтверждают результаты, описанные ранее другими авторами [31][32], указывающими на возможную неэффективность вакцинации в случае недостаточного генетического соответствия между вакцинными и полевыми штаммами вируса ньюкаслской болезни. Обозначенная проблема, в свою очередь, делает актуальной задачу по усовершенствованию мер специфической профилактики ньюкаслской болезни и обновлению вакцинных штаммов в соответствии с эпизоотической ситуацией как в мире, так и в конкретном регионе ее применения.
В результате проведенного опыта было установлено, что все исследуемые вакцины с цельным (неразведенным) содержанием антигена вызывали формирование напряженного поствакцинального иммунитета спустя 28 сут после введения курам, а также соответствовали требованиям ВОЗЖ, предъявляемым к препаратам против ньюкаслской болезни. Тем не менее вакцины на основе штаммов II и VII генотипов вируса ньюкаслской болезни «ВНИИЗЖ-АвиНью Мульти» и «ВНИИЗЖ-АвиНьюФлу Мульти» при контрольном заражении высоковирулентным изолятом вируса ньюкаслской болезни генотипа VII показали более высокий уровень протективной активности при использовании разведенного антигена.
Вклад авторов: Вершинина М. А. – формирование идеи, проведение исследований, анализ и интерпретация полученных данных, принятие ответственности за все аспекты работы, целостность всех частей статьи и за ее окончательный вариант; Мороз Н. В. – формирование идеи, формулировка и развитие ключевых целей и задач; Фролов С. В. – анализ и интерпретация полученных данных, критический пересмотр черновика рукописи с внесением ценных замечаний интеллектуального содержания.
Contribution of the authors: Vershinina M. A. – conceptualization, investigation, data analysis and interpretation, and supervision, project administration, and responsibility for final manuscript; Moroz N. V. – conceptualization, formulation and development of key aims and objectives; Frolov S. V. – data analysis and interpretation, critical revision of the manuscript draft with valuable intellectual input.
1. https://docs.cntd.ru/document/728138234
2. Newcastle disease (infection with Newcastle disease virus). In: WOAH. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.3.10. https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.10_NEWCASTLE_DIS.pdf
3. Сосипаторова В. Ю., Чвала Ир. А., Циванюк М. А., Алтунин Д. А., Чвала Ил. А. Методические рекомендации по идентификации вирусов гриппа птиц и ньюкаслской болезни в реакции торможения гемагглютинации: № 27-16. Владимир: ФГБУ «ВНИИЗЖ»; 2016. 15 с.
4. https://docs.cntd.ru/document/1200158776
5. Newcastle disease (infection with Newcastle disease virus). In: WOAH. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.3.10. https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.10_NEWCASTLE_DIS.pdf
1. Shakal M., Maher M., Metwally A. S., AbdelSabour M. A., Madbbouly Y. M., Safwat G. Molecular identification of a velogenic Newcastle disease virus strain isolated from Egypt. Journal of World’s Poultry Research. 2020; 10 (2S): 195–202. https://doi.org/10.36380/jwpr.2020.25
2. Xiang B., Chen L., Cai J., Liang J., Lin Q., Xu C., et al. Insights into genomic epidemiology, evolution, and transmission dynamics of genotype VII of class II Newcastle disease virus in China. Pathogens. 2020; 9 (10):837. https://doi.org/10.3390/pathogens9100837
3. Liu H., Wang Z., Wang Y. The history and current status of Newcastle disease. China Animal Health Inspection. 2015; (6): 1–4. https://doi.org/10.3969/j.issn.1005-944X.2015.06.001 (in Chinese)
4. Bahoussi A. N., Shah P. T., Zhao J.-Q., Wang P.-H., Guo Y.-Y., Wu C., Xing L. Multiple potential recombination events among Newcastle disease virus genomes in China between 1946 and 2020. Frontiers in Veterinary Science. 2023; 10:1136855. https://doi.org/10.3389/fvets.2023.1136855
5. Roberts J. R., Souillard R., Bertin J. 16 – Avian diseases which affect egg production and quality. In: Improving the Safety and Quality of Eggs and Egg Products. Ed by Y. Nys, M. Bain, F. V. Immerseel. Woodhead Publishing; 2011; 376–393. https://doi.org/10.1533/9780857093912.3.376
6. Absalón A. E., Cortés-Espinosa D. V., Lucio E., Miller P. J., Afonso C. L. Epidemiology, control, and prevention of Newcastle disease in endemic regions: Latin America. Tropical Animal Health and Production. 2019; 51 (5): 1033–1048. https://doi.org/10.1007/s11250-019-01843-z
7. Newcastle Disease, Other Avian Paramyxoviruses, and Avian Metapneumovirus Infections. In: Diseases of Poultry. Ed by D E. Swayne. 13th ed. Ames: Wiley-Blackwell; 2013; Chapter 3: 87–138. https://doi.org/10.1002/9781119421481.ch3
8. Dortmans J. C. F. M. Virulentie Determinanten van het Newcastle Disease Virus: Proefschrift ter verkrijging van de graad van doctor. Utrecht: Universiteit Utrecht; 2011. 134 p. (in het Nederlands)
9. Wang X., Gong Z., Zhao L., Wang J., Sun G., Liu Y., et al. Complete genome sequences of Newcastle disease virus strains isolated from three different poultry species in China. Genome Announcements. 2013; 1 (4):e00198-12. https://doi.org/10.1128/genomea.00198-12
10. Miller P. J., Decanini E. L., Afonso C. L. Newcastle disease: evolution of genotypes and the related diagnostic challenges. Infection, Genetics and Evolution. 2010; 10 (1): 26–35. https://doi.org/10.1016/j.meegid.2009.09.012
11. Dzogbema K. F.-X., Talaki E., Batawui K. B., Dao B. B. Review on Newcastle disease in poultry. International Journal of Biological and Chemical Sciences. 2021; 15 (2): 773–789. https://doi.org/10.4314/ijbcs.v15i2.29
12. Patel S. S., Chauhan H. C., Kumar Sharma K., Patel A. C., Bulbule N. R., Raval S. H., et al. Genetic evolution of Newcastle disease virus subgenotype VII.2 isolates, diagnosed from vaccinated poultry farms of Gujarat, India. Gene. 2024; 930:148859. https://doi.org/10.1016/j.gene.2024.148859
13. Miller P. J., Afonso C. L., El Attrache J., Dorsey K. M., Courtney S. C., Guo Z., Kapczynski D. R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Developmental & Comparative Immunology. 2013; 41 (4): 505–513. https://doi.org/10.1016/j.dci.2013.06.007
14. Eid A. A. M., Hussein A., Hassanin O., Elbakrey R. M., Daines R., Sadeyen J. R., et al. Newcastle disease genotype VII prevalence in poultry and wild birds in Egypt. Viruses. 2022; 14 (10):2244. https://doi.org/10.3390/v14102244
15. Dimitrov K. M., Ramey A. M., Qiu X., Bahl J., Afonso C. L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infection, Genetics and Evolution. 2016; 39: 22–34. https://doi.org/10.1016/j.meegid.2016.01.008
16. Mihiretu B. D., Usui T., Chibssa T. R., Yamaguchi T. Genetic and antigenic characteristics of genotype VII.1.1 Newcastle disease viruses currently circulating in Ethiopian chickens. Virology Journal. 2025; 22 (1):63. https://doi.org/10.1186/s12985-025-02686-x
17. Bello M. B., Yusoff K., Ideris A., Hair-Bejo M., Peeters B. P. H., Omar A. R. Diagnostic and vaccination approaches for Newcastle disease virus in poultry:the current and emerging perspectives. BioMed Research International. 2018; 2018 (1):7278459. https://doi.org/10.1155/2018/7278459
18. Sabouri F., Vasfi Marandi M., Bashashati M. Characterization of a novel VIIl sub-genotype of Newcastle disease virus circulating in Iran. Avian Pathology. 2018; 47 (1): 90–99. https://doi.org/10.1080/03079457.2017.1376735
19. Dimitrov K. Newcastle Disease in Poultry (Avian Pneumoencephalitis, Exotic Newcastle Disease). MSD Veterinary Manual. https://www.msdvetmanual.com/poultry/newcastle-disease-and-other-paramyxovirusinfections/newcastle-disease-in-poultry
20. Meher M. M., Islam J., Afrin M. Investigation of risk factors and biosecurity measures associated with prevalence of Newcastle disease virus in broiler farms. Turkish Journal of Agriculture – Food Science and Technology. 2020; 8 (11): 2426–2432. https://doi.org/10.24925/turjaf.v8i11.2426-2432.3710
21. Hassanzadeh M., Abedi M., Bashashati M., Yousefi A. R., Abdoshah M., Mirzaie S. Evaluation of the Newcastle disease virus genotype VII-mismatched vaccines in SPF chickens: a challenge efficacy study. Veterinary and Animal Science. 2024; 24:100348. https://doi.org/10.1016/j.vas.2024.100348
22. Xu X., Ding Z., Yuan Q., Ding J., Li J., Wang W., et al. A genotype VII Newcastle disease virus-like particles confer full protection with reduced virus load and decreased virus shedding. Vaccine. 2019; 37 (3): 444–451. https://doi.org/10.1016/j.vaccine.2018.11.068
23. Moustapha A., Talaki E., Akourki A., Ousseini M. Newcastle disease virus in poultry: current status and control prospects. World’s Veterinary Journal. 2023; 13 (2): 240–249. https://doi.org/10.54203/scil.2023.wvj26
24. Hu Z., Hu S., Meng C., Wang X., Zhu J., Liu X. Generation of a genotype VII Newcastle disease virus vaccine candidate with high yield in embryonated chicken eggs. Avian Diseases Digest. 2011; 55 (3): 391–397. https://doi.org/10.1637/9633-122410-reg.1
25. Sultan H. A., Elfeil W. K., Nour A. A., Tantawy L., Kamel E. G., Eed E. M., et al. Efficacy of the Newcastle disease virus genotype VII.1.1-matched vaccines in commercial broilers. Vaccines. 2022; 10 (1):29. https://doi.org/10.3390/vaccines10010029
26. Dewidar A. A. A., Kilany W. H., El-Sawah A. A., Shany S. A. S., Dah- shan A.-H. M., Hisham I., et al. Genotype VII.1.1-based Newcastle disease virus vaccines afford better protection against field isolates in commercial broiler chickens. Animals. 2022; 12 (13):1696. https://doi.org/10.3390/ani12131696
27. Фёрстер Э., Рёнц Б. Методы корреляционного и регрессионного анализа: руководство для экономистов. М.: Финансы и статистика; 1983. 304 с.
28. Ван дер Варден Б. Л. Математическая статистика. М.: Издательство иностранной литературы; 1960. 434 с.
29. Урбах В. Ю. Статистический анализ в биологических и медицинских исследованиях. М.: Медицина; 1975. 297 с.
30. Прозоровский В. Б. Статистическая обработка результатов фармакологических исследований. Психофармакология и биологическая наркология. 2007; 7 (3–4): 2090–2120. https://elibrary.ru/jvwcbj
31. Kapczynski D. R., King D. J. Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastle disease outbreak. Vaccine. 2005; 23 (26): 3424–3433. https://doi.org/10.1016/j.vaccine.2005.01.140
32. Miller P. J., King D. J., Afonso C. L., Suarez D. L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine. 2007; 25 (41): 7238–7246. https://doi.org/10.1016/j.vaccine.2007.07.017
Вершинина Мария Андреевна, аспирант, специалист лаборатории профилактики болезней птиц
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Мороз Наталья Владимировна, канд. вет. наук, заведующий лабораторией профилактики болезней птиц
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Фролов Сергей Владимирович, канд. вет. наук, начальник отдела профилактики болезней птиц
ул. Гвардейская, 6, мкр. Юрьевец, г. Владимир, 600901
Вершинина М.А., Мороз Н.В., Фролов С.В. Оценка иммуногенного действия вакцин против ньюкаслской болезни. Ветеринария сегодня. 2026;15(2):177-183. https://doi.org/10.29326/2304-196X-2026-15-2-177-183
Vershinina M.A., Moroz N.V., Frolov S.V. Assessment of immunogenic activity of Newcastle disease vaccines. Veterinary Science Today. 2026;15(2):177-183. https://doi.org/10.29326/2304-196X-2026-15-2-177-183
600901, Владимирская область, город Владимир, микрорайон Юрьевец, улица Гвардейская, дом 6
ФГБУ «ВНИИЗЖ»
тел.: 8 (4922) 26-15-12, доб. 22-27
E-mail: nikeshina@arriah.ru






























